
PRA NÃO DIZER QUE NÃO FALEI DAS FLORES E DO CRIACIONISMO - GERALDO SODRÉ
Estas flores parecem ser uma zueirinha de Deus com os macacos, ou com o darwinismo ? Foram brincadeiras genéticas daquele que montou o complexo sistema do DNA (já super complexo nas camadas cambrianas) , ou estas semelhanças morfológicas indicam parentesco distante entre macacos e estas engraçadas flores (atavismo)?
Seguidores do darwinismo parecem não ter limites em relacionar parentescos infindos por quaisquer semelhanças, declaram que quanto mais diferentes são os seres vivos , mais antigo é o parentesco .
Para evidenciar isso selecionam peças de diferentes seres vivos para quantificar semelhanças e grau de parentesco .
Na natureza "onde nada se cria e apenas se transforma", seres vivos para poderem acontecer teriam que ser criados e quatrilhões de aspectos de engenharias , complexidades , sistemas e informações, demonstrarem que foram serem nitdamente criados por um mesmo padrão genético e portanto um mesmo codificador e fabricante, pois o fato de fazermos, com toda tecnologia genética , comparativamente, menos que nada, mesmo conhecendo genética e podendo manipulá-la para resolver , com imensa dificuldade tecnológica, apenas algumas coisinhas; já deveria a muito tempo nos convencer que se laboratórios tem infinito limite para trabalhar naquilo que já existe, um trilhão de vezes mais teria teorias especulativas de que o nada ou acaso, espontaneamente, teriam o "poder" de, por meio do caos inorgânico racêmico, superar com a seleção natural cega , até mesmo o pior laboratório genético.
Ainda "bem" que a " ciência" agora ensina que uma trombada de um meteoro trazendo super DNA (panspermia) resolveria mais esta "teoria" da vida ..menos mal.
Devemos considerar também que meras semelhanças não indicam necessariamente parentesco filogenético. Elas podem muito mais indicar mesmo fabricante usando os mesmos legos (genes) . Vamos analisar essas questões polêmicas e inseguras ensinadas dogmaticamente com dinheiro público?
Semelhança de alguma coisa não pode ser nunca prova suficiente de ancestralidade comum, pois existe uma DESCONTINUIDADE de grupos em torno de familias de espécies na biodiversidade de HOJE que reflete o mesmo PADRÃO fóssil, que tambem é caracterizado por descontinuidade em torno de familias. Ou seja, a descontinuidade é fato tanto nos fósseis quanto na nossa biodiversidade hoje.
Ocorre tambem outro dado interessante: O mesmo número (quase o mesmo) de arquétipos e modelos de seres vivos nos fósseis com 250 a 300 mil especies, quando comparado ao NÚMERO de arquétipos e modelos esquemáticos de seres vivos hoje nas 2 a 10 milhões de especies, expressadas em bilhões de formas variadas, "é praticamente o mesmo" ( Recebi esta resposta num e-mail recebido de uma das maiores autoridades em estatística fóssil, o geólogo com doutorado em Harvard , Dr Kurt Patrick Wise, que defendeu sua tese de 1200 páginas sobre estatística fóssil , orientado por nada mais nada menos que Stephen Jay Gould, pai do pontualismo paleontológico).
Isto revela claramente que a biodiversidade atual NÃO PASSA de variações sub-especiativas advindas dos, relativamente, poucos tipos básicos ancestrais fósseis; Por isso destaquei mesmo NÚMERO de arquétipos e modelos esquemáticos de seres vivos. Nada se cria na natureza e tudo se transforma dentro dos mesmos limites taxonômicos fósseis até hoje.
Outra prova correlacionada muito contundente, é o fato destes modelos de seres vivos surgirem PRONTOS no registro fóssil (sem etapas evolutivas em camadas anteriores) e isso portanto revela surgimento rápido , aparecimento repentino, o que evoca a idéia de criação deles e não surgimento evolutivo ALGUM .
Esse surgimento de modelos prontos no registro fóssil, tambem evoca a idéia testada e publicada por dezenas de bioquímicos, alegando haver necessidade de que o todo de várias peças do sistema existisse para que houvesse sobrevivência (complexidade irredutível - Behe, Axel, Meyer, Eberlin, Loening, Sanford, etc..
http://www.sensortime.com/loennig-dygmosoic-e.htm)
Se não bastasse isso, ainda temos o fato de que a morfologia dos fósseis se repete em 71%, uma verdadeira LADAINHA DE FORMAS FÓSSEIS; repeteco, e este tipo de amostragem , estatisticamente, só pode se referir a sepultamento de várias populações, pois as espécies são plásticas, mutantes, ultra sensíveis as mudanças ambientais e portanto nunca se repetiriam morfologicamente tanto; nunca, never, jamé.
As proprias espécies que surgem prontas no registro fóssil são plásticas (Herbert citado por Darwin) e vão gastando recursos adaptativos que superam dificuldades de natureza genética e ambiental.
Ou seja, não existiu historia da evoluçao alguma, existiu apenas sobrevivência adaptativa plástica do que sobreviveu.
O que chamamos de evolução, é apenas variação adaptativa sobrevivente de formas de vida , em média, cada vez mais desgastadas e doentes (entropia genética) derivadas de tipos basicos ancestrais mais saudáveis, em geral maiores, mais longevos e mais belos e coloridos (pois cores opacas e sem muita graça, na violencia tanto para caçar e ser visto, como para se esconder, sobrevivem mais ).
No Brasil militarizado, Geraldo Vandré em 1968, ao perceber a falência da ditadura militar em administrar, compos o hino da libertação " Pra não dizer que não falei das flores" onde conclamava a multidão de brasileiros dizendo: " vem vamos embora que esperar não é saber, quem sabe faz a hora não espera acontecer". Hoje, nós criacionistas cantamos as mesmas coisas pra um exército de pensadores, todos prontos com suas armas para impedir a mudança de paradigma educacional, para que possamos trazer novamente o Criador ao seu trono no mundo acadêmico e ao coração da nossa juventude estudantil tão carente de ciência e de Deus .
(2017, Sodre GB Neto )
Artigo
https://www.academia.edu/39120976/Speciation_in_real_time_and_historical-archaeological_and_its_absence_in_geological_time?email_work_card=view-paper
Leitura complementar
http://www.sensortime.com/loennig-dygmosoic-e.htm
https://www.researchgate.net/project/IMPACT-NEOCASTROPHYSIS-THEORY/update/59951859b53d2ff30bda43c0
Canal "Me Salva do Ateísmo"
https://youtu.be/NbeJGS08uC4
Datação e Origem da Radioatividade
http://www.creationscience.com/onlinebook/Radioactivity2.html#wp7826136
http://entropia.comunidades.net/teoria-do-neocatastrofismo-de-impactos-sodre-neto
minha linha de pesquisa questiona datação mais ou menos assim https://www.youtube.com/watch?v=WM_vFJOigj4&t=722s
Dinossauros e homens na arqueologia e historia
http://s8int.com/phile/dinolit87.html
Link contendo 200 referências científicas
http://entropia.comunidades.net/teoria-do-neocatastrofismo-de-impactos-sodre-neto
Link de Walter Brown - Origem da Radioatividade
http://www.creationscience.com/onlinebook/Radioactivity2.html#wp7826136
Link da Publicação de John Baumgardner divulgando testes C14
I address these issues in my chapter, Chapter 8 14C Evidence for a Recent Global Flood and a Young Earth, in the book Radioisotopes and the Age of the Earth: Results of a Young-Earth Creationist Research Initiative. All the chapters of this book are posted online at http://www.icr.org/rate2. Discussion of the diamond results are in section 6. of this chapter 8. I hope this is helpful. John
https://pt.quora.com/Como-pode-um-ser-passar-pela-terra-a-150-milh%C3%B5es-de-anos-e-ainda-ter-vest%C3%ADgio-desse-ser-na-terra-como-no-caso-dos-dinossauros/answer/Sodr%C3%A9-Gon%C3%A7alves-de-Brito-Neto?ch=99&share=583069e4&srid=ZZ5ln
Para TNI - Teoria Neocatastrofista de Impactos clique AQUI
Especiação em tempo real e histórico-arqueológico e sua ausência em tempo geológico (em inglês)
Sodré Gonçalves de Brito Neto1, Everton Fernando Alves2*
1Graduando em Geologia, Universidade Federal de Goiás, UFG, Brasil.
2Departamento de Biotecnologia, Genética e Biologia Celular, Universidade estadual de Maringá, UEM, Paraná, Brasil.
RESUMO
Durante décadas a Biologia evolutiva tem feitos esforços para entender o significado de “espécies” e explicar o seu processo de formação. Atualmente, existem mais de vinte conceitos diferentes de espécies. O uso de diferentes conceitos leva a comparações impróprias e equivocadas. De outro lado, biólogos catastrofistas por décadas têm utilizado o termo “tipo” ou “grupo” para o que consideram categorias de organismos não relacionados geneticamente. Assim, cada uma das várias categorias de espécies, subespécies e variedades que vemos hoje foram concebidas para se diversificarem de um tipo básico ancestral comum. Padrões morfológicos em torno do táxon gênero se identificam com fósseis ancestrais básicos sepultados num catastrofismo recente, e este modelo nos conta uma história de um período de: 1) especiação rápida, 2) permanência com alto número de espécies em ambiente estável (repetição fóssil sem pressões ambientais evolutivas), 3) desastre soterrador de população viva evidenciada pelas repetições de mesmas espécies fósseis (o que descaracteriza a pontualidade), 4) presença de diversas espécies diferentes unidas no registro fóssil, fósseis de imensos vertebrados completos (o que caracteriza desastre de grande magnitude e altas taxas de sedimentação), e 5) mudança drástica no ambiente gerando a radiação das espécies nas camadas recentes de forma amostral e nas milhões de espécies na biodiversidade atual.
Palavras-chave: Tipos básicos ancestrais, catrastrofismo, especiação rápida, estase morfológica, pontualismo.
INTRODUÇÃO
Nos últimos séculos, a biologia evolutiva tem feito esforços para entender o significado de “espécies” e explicar o seu processo de formação. Atualmente, existem mais de vinte conceitos diferentes de espécies (de Queiroz, 2005). O uso de diferentes conceitos leva a comparações sob critérios distintos, contudo tal indefinição na ponta taxonômica (táxon espécie) nos leva a definir em grandezas taxonômicas mais abrangentes. Neste trabalho, consideraremos a grandeza PTMG (Padrões morfológicos em torno do táxon gênero) como referência.
Muitas das espécies e suas variações, observáveis hoje, refletem os mesmos padrões morfológicos em torno do táxon pertencente ao nível de gênero. Padrões morfológicos em torno do táxon gênero (PMTG) são, portanto, o que consideramos como as características peculiares dos tipos básicos matrizes, que ainda hoje estão presentes na maioria das diversas espécies e suas variações encontradas na natureza (Eldredge e Stanley, 1984). Muitas espécies viventes hoje, e suas variações, refletem padrões morfológicos semelhantes. A permanência destes mesmos padrões morfológicos fósseis, por mais que aumente o número de espécies, nos estimula a formularmos perguntas importantes a serem consideradas aqui.
A tese dos tipos básicos matrizes ou ancestrais (Marsh, 1941), tem sido apoiada pelo fato de o registro fóssil apresentar baixa variabilidade (estase morfológica) e baixa radiação especiativa entre as espécies fósseis (Zimmerman, 1960; Martens, 1997; Albrecht e Wilke, 2008; Alisson, 2013), independente da suas respectivas plasticidades ou maleabilidades fenotípicas ou genotípicas (Ghalambor et al., 2015). Além disso, outros fatores que corroboram a tese dos tipos básicos ancestrais são os mais de 4229 gêneros bem documentados de “fósseis vivos” que são chamados assim por terem sofrido poucas mudanças ao longo do tempo, logo, mantendo-se semelhantes às encontradas no registro fóssil (Romer, 1966; Whitmore, 2013a; Whitmore, 2013b). Provavelmente refletindo idéias de sua época, Darwin chegou a citar essa dedução corrente ao escrever:
"Não há uma verdadeira grandeza nesta forma de considerar a vida, com os seus poderes diversos atribuídos primitivamente pelo Criador a um pequeno número de formas, ou mesmo a uma só? Ora, enquanto que o nosso planeta, obedecendo à lei fixa da gravitação, continua a girar na sua órbita, uma quantidade infinita de belas e admiráveis formas, saídas de um começo tão simples, não têm cessado de se desenvolver e desenvolvem-se ainda!... O venerável e reverendo W. Herbert, mais tarde deão de Manchester, escrevia em 1822, no 4., volume das Horticultural Transactions, e na sua obra as Amaryllidacées (1837, p. 19, 339), que ‘as experiências de horticultura têm estabelecido, sem refutação possível, que as espécies botânicas não são mais que uma classe superior de variedades mais permanentes’. Aplica a mesma opinião aos animais e vê que as espécies únicas de cada gênero foram criadas num estado primitivo muito plástico, e que estes tipos produziram ulteriormente, principalmente pelo cruzamento e também por variação, todas as nossas espécies existentes. Na Nouvelle Flore de l’Amérique Du Nord (1836, p. 6), Rafinesque exprimia-se assim: ‘Todas as espécies podiam ser outrora variedades, e muitas variedades tornaram-se gradualmente espécies, adquirindo caracteres permanentes e particulares’; e um pouco mais adiante (pág. 18) acrescenta: ‘excetuando os tipos primitivos ou ancestrais do gênero’”(Darwin, 1866, capítulo 9, pp.25-26, 577, ênfase nossa).
Quando falamos de estase morfológica no registro fóssil, nos referimos às camadas sedimentares compreendidas entre o Ediacarano/Cambriano até próximo das camadas do período Pleistoceno, como demonstra diversas publicações baseadas na teoria do equilíbrio pontuado (pontualismo) proposta pelos paleontólogos NilesEldredge e Stephen Gould (Levinton e Chris, 1980; Woodruff, 1980; Williamson, 1981; Eldredge, 1986; Van Bocxlaer e Hunt, 2013). E não nas camadas atuais onde a variação irradia em multiformes variações.
Os padrões morfológicos ligados ao táxon gênero podem ser observados no registro fóssil surgindo repentinamente em todas as camadas sedimentares, com expressivo aparecimento no cambriano e surgimentos abruptos sem nenhum gradualismo em toda coluna geológica, mas uma repetição surpreendente. Essa constatação tem sido divulgada há décadas pelos adeptos do pontualismo que admitem a estase morfológica e a radiação florescendo em especiações somente nas camadas geológicas mais atuais (Levinton e Chris, 1980; Woodruff, 1980; Williamson, 1981; Eldredge, 1986; Van Bocxlaer e Hunt, 2013).
Estudo evolutivo recente corrobora tal afirmação ao dizer que
"a visão dominante da evolução baseada no registro fóssil é que as espécies estabelecidas permanecem mais ou menos inalteradas durante sua existência. Por outro lado, a evolução substancial é rotineiramente relatada em populações contemporâneas, e a maioria dos traços quantitativos mostra um alto potencial de evolução. Essas observações contrastantes em escalas de tempo longo e curto são muitas vezes referidas como o paradoxo da estase, que se baseia na suposição fundamental de que os períodos de estase morfológica no registro fóssil representam uma mudança evolutiva mínima." (Voje, 2016).
Assim, o presente artigo tem como objeto de estudo esses três fatos observados e bem documentados: surgimento repentino fóssil, estase morfológica e repetição de mesmas espécies fósseis e explosão de radiação adaptativa revelando todo potencial de plasticidade e maleabilidade dos fenótipos e genótipos dos seres vivos.
As especiações e variações que, como vimos, carregam os PMTG podem ser observadas ocorrendo em tempo real e histórico-arqueológico, mas estão ausentes na mesma proporção, no registro fóssil. Isto se torna relevante na medida em que percebemos o sucesso reprodutivo das variações e sua permanência ao longo do tempo, pois a variação em torno de gênero não exige grandes bio-transformações, mas deve ocorrer de qualquer maneira nas amostras fósseis essas variabilizações que estão cada vez mais permanente e cada vez mais com sucesso reprodutivo (com exceção das extintas). Hoje, surpreendentemente, temos quase o mesmo número de padrões corporais entre os cerca de 2 milhões de espécies vivas já catalogadas (9 a 100 milhões estimadas) e as ínfimas 300 mil espécies fósseis do cambriano/edicara ao pleistosceno. (Woodmorappe, 2000; Sadava et al., 2009; Mora et al., 2011; Catalogue of Life, 2016). Quando comparados os padrões morfológicos, esperar-se-ia que tivéssemos um número bem maior de PMTG no registro fóssil se este espelhasse uma amostra da biodiversidade ocorrida em 544 milhões de anos correspondentes ao Fanerozóico. Portanto, o fato do número de espécies atuais, na casa dos milhões, não conseguir transpor o número de padrões corporais presente no registro fóssil, sugere evolução delimitada a padrões morfológicos ou tipos básicos ancestrais (baraminologia) ou PMTG.
Subtipos de especiação rápida
Entendemos a especiação em tempo real como um fenômeno em que duas ou mais populações de uma mesma espécie se modificam em novos arranjos de informação genética pré-existente, tendo como causa a separação por barreiras geográficas e mortandades, porém em um tempo hábil para que seja possível a observação de todo o processo do início ao fim.
Podemos ainda classificar o processo de especiação de duas formas: em tempo real ou em tempo histórico-arqueológico. A especiação em tempo real é aquela em que ocorre bio-modificações limitadas ao tipo básico, equivalentes ao surgimento rápido de uma nova “espécie”, observada ̶ por experimentação ou observação não-controlada ̶ em até 50 anos. A especiação em tempo histórico, por sua vez, diz respeito às bio-modificações equivalentes a uma nova “espécie” acima de 50 anos que podem ou não ser observadas devido o tempo de vida do observador ou projeto de pesquisa. Nesses casos, há 50% de chance de observação e os outros 50% restantes dependem do uso da dedução, por exemplo, a partir de análise cromossômica e cálculos de taxas de radiação (Trivedi, 2000).
O papel da deriva genética no processo de especiação
Existem diversos mecanismos envolvidos no processo de especiação (epigenética, deriva genética, seleção natural, forças geomagnéticas no crossover, etc). Devido à especiação estar intimamente relacionada à deriva genética e a conseqüente perda de pool gênico de uma população anterior, estudos do modelo de muitos ancestrais básicos (ASCR) que apostam na especiação em tempo real, se tornam fundamentais para explicar as evidências atuais de limites à adaptação nos diferentes organismos vivos (Bell, 2013).
A deriva genética é um mecanismo que modifica aleatoriamente e de forma repentina – devido a catástrofes ou a isolamentos diversos, por exemplo, − as frequencias alélicas de uma população (Ridley, 2006; Freeman e Herron, 2009). Por outro lado, acredita-se que a seleção natural é direcionada, isto é, elimina muitas mutações deletérias, mutações necessárias se perdem juntas, ignora as mutações neutras e deletérias (principalmente as que se manifestam após período reprodutivo) seleciona os traços sobreviventes e/ou vantajosos pré-existentes ou que comunicaram em face de resposta epigenética ao meio (Sanford, 2014). Porém, como afirmou a bióloga americana Lynn Margulis, “a seleção natural elimina, e talvez mantenha, mas ela não cria” (Teresi, 2011, p. 68). Ademais, devido o fato da deriva genética não fazer distinção entre genes bons e ruins, e a seleção natural falhar também em seu filtro, isso resulta em perda de variação genética, deixando as espécies geneticamente mais pobres e “cada vez mais próximas da extinção”. Em outras palavras, a deriva decepa e empobrece a população de seu “pool gênico” e a seleção natural ajeita mais ou menos o que restou.
Sabe-se que o efeito da deriva é maior quando é menor o tamanho da população, podendo aparecer em diferentes momentos da história da humanidade. Nesse sentido, é possível que a deriva tenha tido um papel fundamental e sobrepujante ao da seleção natural no que diz respeito ao processo de especiação rápida após uma grande catástrofe.
O processo de especiação após Catástrofes
Grandes catástrofes mundiais são as formadoras da maioria das camadas sedimentares presentes no globo, que estão associadas a imensas inundações e destruições em todo globo (Souza Jr, 2008). As conseqüências da mortalidade em massa gerada, stress endogâmico nas espécies sobreviventes e isolamentos geográficos, deram, alem de outros fatores, condições muito favoráveis para uma rápida especiação (Wilmer et al., 2011). O modelo ASCR defende o sepultamento da biodiversidade de todo planeta por estratificação espontânea nas transgressões e regressões marinhas, seguido de especiações rápidas pelos motores evolutivos (Berthault, 1986; Berthault, 1988; Brand e Tang, 1991;Snelling, 1997; Berthault, 1998; Chadwick e Spencer, 2006; Berthault, 2013).
O modelo ASCR de história curta da formação das camadas sedimentares prevê um momento em que os poucos seres humanos e animais sobreviventes e isolados por essas catástrofes associadas (de energia e magnitude global) teriam recomeçado a repovoar a terra, e sucessivas derivas e stress endogâmicos de cruzamentos consangüíneos haveriam de ocorrer, exatamente como percebemos em espécies de Denisovans (Prüfer et al., 2013) e nas fundações de etnias de biotipos semelhantes entre si habitando em cima de fósseis de halogrupos dessemelhantes como, por exemplo, mongolóides nas Américas em cima de fósseis dessemelhantes negroides em Lagoa Santa-MG, São Paulo e México. Fato observado também na China de mongolóides habitando em cima de fósseis caucasianos tanto nas etnias humanas quanto em milhares de outros seres vivos semelhantes entre si isolados de seus parentes.
Nesse contexto, a especiação em tempo real legitima o modelo de um “salto evolutivo” recente, com dois picos bio-diferenciadores relacionados aos episódios do inicio da entropia genética humana (Fu et al., 2013) depois das catástrofes associadas que empobreceriam o pool gênico dos seres vivos e dariam aos sobreviventes, em virtude da migração e do isolamento geográfico, o legado de efeito fundador – situação frequente na especiação peripátrica− em um planeta com um ecossistema totalmente reconfigurado onde o motor epigenético atuaria para corresponder as necessidades do novo ambiente (Ridley, 2006;Eakin, 2014; Weyrich et al., 2016).
A especiação peripátrica é um mecanismo pelo qual podemos explicar o enorme aumento da diversificação pós-catástrofes. É um tipo de especiação pelo qual as espécies novas são formadas em populações periféricas isoladas (Ridley, 2006). Na especiação peripátrica, populações drasticamente reduzidas fazem com que a especiação completa seja o resultado mais provável do isolamento geográfico, porque a deriva genética age mais rapidamente em populações pequenas. Deriva genética somada a fortes pressões seletivas, causariam uma rápida mudança genética na pequena população descendente (Wilmer et al., 2011).
Observações atuais podem servir de exemplo para compreendermos melhor os casos de especiação peripátrica. Estudo recente afirma que “uma análise de mais de 2.000 espécies de pássaros fornece uma visão sobre como evoluíram as diversas formas de bico dos animais e aponta para um único evento raro como um gatilho para a rápida divergência inicial das linhagens aviárias.” (Bhullar, 2017).
Toda essa sucessão de fatos pode ser vislumbrada por meio de um cenário onde a maioria dos organismos destruídos por essas grandes catástrofes associadas e consequentes, deixaria pequenas populações de sobreviventes (Wilmer et al., 2011). Segundo matéria publicada pela Folha de São Paulo a respeito desse achado,
“o que pareceu fascinar mais outros biólogos, porém, é a grande velocidade com que o fenômeno do deslocamento de caráter ocorreu. ‘Eu acreditava que fosse demorar muito mais’, comentou na ‘Science’ o biólogo David Pfening, da Universidade da Carolina do Norte. A redução média de 5% no tamanho de bico, considerada drástica pelos biólogos, ocorreu no intervalo de cerca de um ano, praticamente de uma geração para a outra.” (Folha de S. Paulo, 2006).
A surpresa dos biólogos evolutivos com a descoberta desse novo fenômeno se dá devido à falta de entendimento de outros motores coadjuvantes, que atuam nessas bio-modificações limitadas ao tipo básico, tais como epigenética e aspectos energéticos e de temperatura atuando no crossover (Fondon e Garner, 2004; Eakin, 2014). Para a Dra. Jean K. Lightner,45 parece haver três fontes para as variações associadas a radiação adaptativa: 1) hibridização, 2) mutação e 3) triagem ambiental de alelos ancestrais (por mecanismos de seleção natural e deriva meiótica) (Lightner, 2016).
Estas observações, se não encarecem a dívida dos elos perdidos na paleontologia pontualista, aumenta muito a dívida de variabilização taxonômica fóssil para o modelo histórico geológico que assume pelo menos cinco grandes catástrofes separadas por milhões de anos na história da terra.
O curioso é que as pesquisas ligadas a especiação em tempo real tanto confirmam as observações de Darwin no âmbito da biologia funcional (equilíbrio pontuado) quanto destrói por completo os postulados evolutivos em termos de períodos geológicos (gradualismo filético), e é perfeitamente comunicável ao modelo catastrofista bíblico que aposta numa especiação rápida impulsionada pelo efeito das bio-modificações limitadas ao tipo básico.
Baraminologia e o estudo dos “tipos básicos”
Há uma comunicação muito fluente entre o darwinismo biológico e o modelo ASCR biológico, onde muitos se valem de descobertas evolutivas, como especiação em tempo real, para se aproximar mais da historicidade de curto período de tempo, e utilizando a radiação recente como justificativa para toda a biodiversidade. A teoria da evolução sintética, mais especificamente atuando no âmbito biológico, que ensina a diferenciação das espécies, neste contexto, tem baixa discórdia entre as classes mais avançadas de ASCR, sobretudo nos cientistas do movimento de “Baraminologia” (Marsh, 1941; Wise, 1992; Robinson e Cavanaugh, 1998; Frair, 2000; Jerlström, 2000; Wood e Cavanaugh, 2001; Cavanaugh e Wood, 2002; Wood, 2010; Aaron, 2014). Suas discordâncias se concentram mais no âmbito da história geo-paleontológica compreendida entre o cambriano/edicara e pleistoceno.
Desde 1941, o modelo ASCR é contrário a ideia de que especiação é sinônimo de “evolução” por meio de estudos relacionados às especiações no campo da Baraminologia (Marsh, 1941; Wise, 1992; Robinson e Cavanaugh, 1998; Frair, 2000; Jerlström, 2000; Wood e Cavanaugh, 2001; Cavanaugh e Wood, 2002; Wood, 2010; Aaron, 2014). O biólogo norte-americano Dr. Frank L. Marsh, um dos fundadores da Creation Research Society foi quem cunhou a palavra "baramin". (Marsh, 1941; Frair, 2000) Ela foi derivada da combinação de duas palavras hebraicas − bara (criado) e min (tipo) – referindo-se a tipos básicos criados (espécies, em versões bíblicas em português) (Frair, 2000).
Em 1990, o paleontólogo catastrofista Kurt Wise observou a necessidade de uma biossistemática ASCR − um método de estudo, nomeação, e classificação de baramin (Wise, 1990; Frair, 2000) ou ancestrais de PTMG como defendemos aqui. O campo científico foi oficialmente denominado como “baraminologia”, que de forma simplista significa o estudo dos baramins ou dos tipos básicos ancestrais. Segundo os pesquisadores Reinhard Junker e Siegfried Scherer,“tipos básicos é uma unidade de classificação, um taxon, resultado do trabalho da descontinuidade sistemática como é observado na natureza” (1996, p.34; Wood et al., 2003). Dito de forma simples, tipos básicos criados variabilizaram ao longo do tempo até chegarem ao que conhecemos hoje como subespécies.
Existem algumas regras falseáveis para se considerar um grupo de espécies como pertencentes a um tipo básico ancestral comum. O Dr. Junker e o Dr. Siegrifield Scherer destacam na 6ª edição alemã do livro Evolução, um livro texto crítico:
“todos os indivíduos que estão unidos direta ou indiretamente por cruzamentos são considerados pertencentes a um tipo básico (nível genético). E todas as espécies biológicas que se se assemelham claramente umas as outras pertencem a um gênero (nível morfológico). E todas as espécies biológicas que em princípio podem cruzar entre si pertencem a um tipo básico (nível morfo-genético)” (Junker e Scherer, 1996, p.34).
Os autores ainda acrescentam que “dois indivíduos pertencem ao mesmo tipo básico quando a embriogênese de um híbrido vai além da fase maternal do desenvolvimento e contem uma expressão coordenada e genes morfogenéticos paternos e maternos” (Junker e Scherer, 1996, p.34). Além disso, os baraminologistas usam uma série de critérios metodológicos de adesão para determinar os limites dos grupos baramins (Robinson, 1997; Wood, 2001; Wood, 2002; Wood, 2005; Cavanaugh e Sternberg, 2004). De modo geral, os métodos mostram espacialmente graus de similaridade e de dissimilaridade entre grupos, e podem revelar informações taxonômicas úteis, distinguindo cada vez mais os fatores que dão probabilidade ou não de parentesco, aumentando assim sua contribuição em biologia aplicada a técnicas de melhoramento genético e estudo do comportamento evolutivo das populações.
A baraminologia, também conhecida como sistemática de descontinuidade, está rapidamente se tornando uma das áreas mais ativas de pesquisa ASCR (Scherer, 1993), e algumas de suas metodologias têm sido aplicadas e testadas até mesmo por pesquisadores ligados ao modelo gradualista geológico de ancestralidade totalmente comum com destaques em periódicos com peer-review (Senter, 2010; Wood, 2011). Como já vimos, sua principal finalidade é determinar quais organismos compartilham um ancestral comum (Frair, 2000). A ideia básica defendida neste campo de pesquisa é a de que há limites nas possibilidades de cruzamento que não podem ser atravessados. Nesse contexto, baraminologistas objetivam encontrar as "descontinuidades" na história de vida, ou os limites da ancestralidade comum (Remine, 1993).
Esse campo de pesquisa ganha ainda mais estímulo frente às evidências atuais mostrando que “especialistas” têm erroneamente classificado algumas espécies dentro de um dado gênero por conta do “desejo” em descobrir o ancestral comum universal (Lopes, 2015). Paleontólogos afirmam que um terço das “espécies” reconhecidas como sendo de dinossauros talvez nem mesmo tenha existido (Horner e Goodwin, 2009). Para eles, essas “espécies” podem não ser espécies separadas, mas estágios juvenis ou subadultos, em desenvolvimento, identificados erroneamente como exemplares de outras espécies. Em artigo publicado na revista Science, por exemplo, Schwartz e Tattersall afirmam que esse milagre da multiplicação da nomenclatura das espécies foi longe demais (Schwartz e Tattersall, 2015).
Vale lembrar que, embora a Baraminologia tenha alcançado resultados promissores, suas conclusões não são definitivas (Wilson, 2010). Por ser um campo recente, mais pesquisas são necessárias e seus métodos e técnicas recém-elaborados devem ser mais bem examinados a fim de legitimar ou não a sua função e utilidade na caixa de ferramentas da ciência.
Entropia genética na especiação
E o que dizer dos custos causados quando realmente ocorre a especiação? Outro grande dificultador para o modelo gradualista é que ele é neutro em relação à melhora ou piora do processo de especiação, embora assuma que a deriva genética cause perdas de variação genética em populações pequenas. O modelo de muitos ancestrais básicos sepultados num catastrofismo recente (ASCR), por sua vez, munido da proposta da entropia genética, defende que a especiação resulta na perda de informação genética e na consequente degeneração do genoma devido às adaptações como recursos evasivos, perigosos, desgastantes para as populações (Ariza, 2007; Sanford, 2014; Crabtree, 2013a; Crabtree, 2013b). Outras evidências corroboram o modelo ASCR no sentido de sugerir que essa perda de informação genética devido a mutações deletérias em humanos tenha se dado recentemente, entre 5.000 a 10.000 anos atrás (Fu, 2013).
A entropia na informação genética é cada vez mais evidente em tudo que se observa: na deriva genética, na seleção/eliminação, nas mutações, nas complexidades imunológicas desgastantes, entre outros mecanismos. Em humanos, por exemplo, as estimativas atuais é a de que ocorram entre 100-200 novas mutações por indivíduo a cada geração (Nachman e Crowell, 2000; Dolgin, 2009; Lynch, 2010). Destas, os dados variam entre 1-15% de mutações deletérias que causariam a perda direta de informação genética em humanos a cada geração (Nachman e Crowell, 2000; Lynch, 2010; Eyre-Walker e Keightley, 1999;Shabalina et AL., 2001; Keightley, 2012).
Em relação ao fitness, um estudo publicado em 1997 estimou entre 1-2% a taxa de perda da aptidão humana, ou seja, um custo genético alto que faz com que a humanidade se degenere a cada geração devido o esgotamento dos recursos adaptativos utilizados para manter sua variedade genética (Crow, 1997). Em 2010, por sua vez, outro estudo estimou que a aptidão humana está em declínio em 3-5% por geração (Lynch, 2010).
O zoólogo holandês Duyvene de Wit descreveu perfeitamente esse processo de empobrecimento genético ao afirmar que:
“quando uma população marginal abre caminho para um novo habitat, ela não pode levar consigo todos os genes da população materna, mas sim apenas parte deles. Cada nova raça ou espécie que se origina de outra possui, portanto, um pool gênico mais pobre. Conseqüentemente, a perda de substância do ‘pool’ gênico é o preço que cada raça ou espécie tem de pagar pelo privilégio de existir. Se o processo de especiação ocorrer repetidas vezes consecutivamente, surgirão por fim, espécies cujos ‘pools’ gênicos estão tão completamente esgotados que bastam alterações insignificantes nas condições ambientais para que elas venham a se extinguir. Esforços para se adaptar a modificações ambientais como resultado de possibilidades insuficientes de recombinação levam, por fim, a um estado genético mínimo. Se este limite mínimo for ultrapassado, não haverá mais possibilidade de sobrevivência. Por essa razão, o destino trágico e irrevogável das espécies ou raças especializadas, extremamente adaptadas, é a morte genética” (Kahle, 1999, p.87; Junker e Scherer, 1996).
Portanto, a proposta ASCR é razoável e se torna cada vez mais corroborada pelos dados científicos ao afirmar que os seres vivos do passado eram mais saudáveis e aptos em relação aos de hoje, sem os defeitos oriundos das adaptações forçadas e das pressões seletivas distintas a fim de que viessem a subsistir. Vale lembrar que sub-especiacões, quanto mais verificáveis em tempo real, mais provam a bio-modificação em curto período de tempo, tal como prevê o modelo catastrofista.
Registro fóssil e a ausência de diversificação de espécies
Especiação proporcional ao comportamento biodiferenciador observado está ausente pelo fato de existir apenas cerca de 300.000 tipos fósseis se repetindo dentre as estimativas de trilhões de amostras, em todos os estratos geológicos até o pleistosceno (Woodmorappe, 2000; Sadava et al., 2009; Loceye Lennon, 2016). Este fato indica a ausência de sequências durante o processo de especiação quando contrastada com as estimativas de 8,7 milhões de espécies vivas atualmente (Mora et al., 2011).
Em outras palavras, a ausência de “diversidade” no registro fóssil revela que as diferenciações ocorrem mais no presente (milhões de espécies) que em toda coluna geológica (centenas de milhares). Este episódio indica um tempo em que os seres vivos não precisavam se sub-especiar (bio-modificar) e se adaptar com frequência pra sobreviver, porque viviam em um ambiente mais propício para a vida.
Evidências recentes apontam que o início da diversificação de alguns gêneros de plantas consideradas “fósseis vivos”, por exemplo, ocorreu ao mesmo tempo ao redor do mundo e em um período muito mais recente do que se supunha, revelando conflitos na proposta gradualista das espécies (Nagalingum et al., 2011). Ademais, foi relatado que essa rápida diversificação teve como causa uma grande mudança climática.
Aliás, são vários os motores adaptacionais e modificacionais integrados reconhecidos atualmente. O fato de eles existirem nos seres vivos, nos permite sugerir que as bio-modificações nunca deixaram de existir. Porém, o curioso é que evolucionistas sugerem que em 90% dos 544 milhões de anos totais do Fanerozóico, as bio-modificações se estagnaram, repetindo na coluna geológica as mesmas espécies, e só recentemente nas camadas mais atuais (superiores) é que houve biodiversificação em larga escala (radiação) (Gould, 1981). A Figura 1 facilita nossa compreensão ao mostrar que em 500 milhões de anos, o gráfico pouco oscila alcançando em média dois milhares de gêneros, ao passo que somente em períodos mais recentes temos a explosão para mais de supostos cinco milhares de gêneros.
Figura 1–A grande explosão de vida no planeta durante o Fanerozóico. Adaptado de Wikipedia (2016).
Dr. Tom Kemp, curador das coleções zoológicas do Museu de História Natural da Universidade de Oxford, por exemplo, fez a seguinte admissão: “como é agora bem conhecido, a maioria das espécies fósseis aparecem instantaneamente no registro fóssil, persistem por alguns milhões de anos praticamente inalterados, e apenas desaparecem abruptamente” (Kemp, 1985, p. 67).
O mesmo fato é observado por Peter Williamson, professor de Geologia da Universidade de Harvard ao sugerir que o neodarwinismo tem falhado em não conseguir explicar as descontinuidades sistemáticas no registro fóssil:
"o principal problema é a estase morfológica. Uma teoria é somente tão boa quanto o são suas predições, e o neodarwinismo convencional, que alega ser uma explicação abrangente do processo evolutivo, falhou por não predizer a ampla estase morfológica, agora reconhecida como um dos aspectos mais impressionantes do registro fóssil." (Williamson, 1981, p.214).
Darwin sugeriu que os seres vivos surgiam por evolução gradativa, e esperava que um dia o registro fóssil confirmasse sua predição, porém não foi o que ocorreu: Darwin estava errado. Dia após dia, os inúmeros fósseis escavados por todo o mundo têm refutado a hipótese de mudança gradual em camadas sobrepostas supostamente cronológicas. Ao contrário do que Darwin esperava, os dados recentes revelam padrões de aparecimento súbito (explosões) seguido por longos períodos de pouca mudança.
O famoso paleontólogo evolucionista Niles Eldredge admitiu na revista New Scientist que:
"os paleontólogos desde Darwin têm feito buscas (em grande parte em vão) de sequências de séries graduadas de fósseis que se destacam como exemplos do tipo de transformação global de espécies que Darwin imaginou como o produto natural do processo evolutivo. Poucos viram qualquer razão para duvidar − embora seja um fato surpreendente que [...] a maioria das espécies permanecem reconhecidamente as mesmas, praticamente inalteradas ao longo de sua ocorrência em estratos geológicos de várias idades" (Eldredge, 1986, p.55).
A ausência de fósseis intermediários é demasiado óbvia para os evolucionistas encobrirem por mais tempo. O biólogo evolutivo Dr. David Woodruff da Universidade da Califórnia expressou na revista Science o desapontamento de evolucionistas relativo à ausência no registro fóssil de formas transicionais: "as espécies fósseis permanecem inalteradas durante a maior parte de sua história e o registro não consegue conter um único exemplo de uma transição significativa." (Woodruff, 1980, p.716).
Portanto, diante de tantas admissões, finalmente concordamos com a bióloga evolucionista Lynn Margulis quando diz que:
“não há nenhum gradualismo no registro fóssil. [...] O ‘equilíbrio pontuado’ foi inventado para descrever a descontinuidade no surgimento de novas espécies. [...] Os críticos, [incluindo os críticos ASCRs], estão certos nas críticas que fazem. [...] Os biólogos evolucionistas acreditam que o padrão evolucionário é uma árvore. Não é!” (Teresi, 2011).
Exemplos de especiação rápida
Existem diversos relatos de surgimento de novas “espécies” em períodos que variam de dezenas há milhares de anos. Segundo essas informações, a especiação é um fenômeno que não necessita de milhões de anos para acontecer. O biólogo norte-americano Dr. James Gibson, Diretor do Instituto de Pesquisa em Geociência (GRI, na sigla em inglês), instituição afiliada à Universidade de Andrews (EUA), mantida pela IASD, descreveu um exemplo de especiação em tempo real: “uma nova espécie de copépode [crustáceos] formou-se no Mar Salton no sul da Califórnia em menos de 30 anos.” (Johnson, 1953; Gibson, 2002).
Estudos diversos têm relatado que apenas entre 10 a 36 anos as populações diferentes de lagartos sofreram alterações morfológicas significativas o suficiente para serem consideradas novas “espécies” (Morrell, 1997;Herrel et al., 2008). Outros exemplos conhecidos de especiação em tempo real incluem bactérias (Shikano et al., 1990), moscas (Huey et al., 2000), tentilhões (Grant e Grant, 2006), rãs (Hoskin et al., 2005), escaravelhos (Halliburton e Gall, 1981) e plantas (Groves e Groves, 1880; Foucaud, 1897; Marchant, 1963).
Dr. Gibson forneceu ainda exemplos de especiação em tempo histórico-arqueológico: “uma população de macacos verdes viveu na ilha de St Kitts no Caribe por menos de 100 anos, mas desenvolveu aspectos morfológicos equivalentes a uma nova espécie.” (Ashton et al., 1979; Gibson, 2002). “O Havaí não tinha bananeiras até cerca de 1000 anos atrás, no entanto há mariposas havaianas nativas que só se alimentam de bananeiras. Estas novas espécies surgiram em menos de 1000 anos.” (Zimmerman, 1960; Gibson, 2002). Diante desses exemplos, Dr. Gibson conclui: “a capacidade de mudanças rápidas está confirmada tanto por experimentação como por observação da natureza” (Gibson, 2002).
Conclusão
Padrões morfológicos em torno do táxon gênero (PMTG) se identificam com fósseis ancestrais básicos sepultados num catastrofismo recente (ASCR), pois este modelo respeita o fato da especiação rápida observada, que exigiria uma variabilidade proporcional muito grande no registro fóssil, caso representasse a evolução em milhões de anos, e isto além de não existir tal variação taxonômica, ainda se confirma com estase morfológica, repetição das mesmas espécies e 4229 gêneros de fósseis vivos, quadro este que reflete um sepultamento repentino dos seres vivos do planeta e não uma suposta história evolutiva da vida. Relatos de especiação na história e arqueologia, capazes de justificar toda biodiversidade em pouco tempo, nos falam também que deveríamos ter muito mais variabilidade no registro fóssil do que há. A explosão cambriana, a ausência de variabilidade amostral fóssil (estase morfológica fóssil) e falta de radiação especiativa no registro fóssil até o pleistoceno, nos conta uma história de um período de: 1) surgimento rápido, 2) permanência com alto número de espécies em ambiente estável (repetição fóssil sem pressões ambientais evolutivas), 3) desastre soterrador de população viva evidenciada pelas repetições de mesmas espécies fósseis (o que descaracteriza a pontualidade), 4) presença de diversas espécies diferentes unidas no registro fóssil, fósseis de imensos vertebrados completos (o que caracteriza desastre de grande magnitude e altas taxas de sedimentação), e 5) mudança drástica no ambiente gerando a radiação das espécies nas camadas recentes de forma amostral e nas milhões de espécies na biodiversidade atual.
REFERÊNCIAS
Ghalambor CK, Hoke KL, Ruell EW, Fisher EK, Reznick DN, Hughes KA (2015). Non-adaptive plasticity potentiates rapid adaptive evolution of gene expression in nature. Nature. 525(7569): 372–375.
Souza Jr NN (2008). Uma breve história da terra. Brasília, DF: Sociedade Criacionista Brasileira.
Aaron M (2014). Baraminological Analysis of the Caseidae (Synapsida: Pelycosauria). Journal of Creation Theology and Science Series B: Life Sciences. 4:19-22.
Albrecht C, Wilke T (2008). Ancient Lake Ohrid: biodiversity and evolution. Hydrobiologia. 615(1):103-140.
Alisson E (2013). Arqueologia ajudará a desvendar origem da biodiversidade amazônica. Agência FAPESP. Disponível em: http://agencia.fapesp.br/arqueologia_ajudara_a_desvendar_origem_da_biodiversidade_amazonica/16937/
Ariza LM (2007). Evolution in a Petri Dish. Scientific American. 297(6):36. Disponível em: http://www.scientificamerican.com/article/evolution-in-a-petri-dish/
Ashton EH, Flinn RM, Griffiths RK, Moore WJ (1979). The results of geographic isolation on the teeth and skull of the Green monkey (Cercopithecus aethiopssabaeus) in St. Kitts – a multivariate retrospect. Journal of Zoology. 188(4):533-555.
Bell G (2013). Evolutionary rescue and the limits of adaptation. Philos Trans R Soc Lond B Biol Sci. 368(1610):20120080.
Berthault G (1986). Experiments on lamination of sediments, resulting from a periodic graded-bedding subsequent to deposition—a contribution to the explanation of lamination of various sediments and sedimentary rocks. ComptesRendus De L Academie Sciences, Paris. 303, Série II (17):1569-1574.
Berthault G (1988). Sedimentation of a heterogranular mixture. Experimental lamination in still and running water. ComptesRendus De L Academie Sciences, Paris. 306, Serie II:717-724.
Berthault G (1998). Genesis and historical geology: a personal perspective. CEN Tech. J. 12(2):213-17.
Berthault G (2013). Orogenesis: Cause of Sedimentary Formations. Open Journal of Geology. 3:22-24.
Bhullar BS (2017). Evolution: Catastrophe triggers diversification. Nature. 542 (7641):304-305.
Brand LR, Tang T (1991). Fossil vertebrate footprints in the Coconino Sandstone (Permian) of northern Arizona: Evidence for underwater origin. Geology. 19(12):1201-1204.
Catalogue of Life (2016). Species 2000 & ITIS Catalogue of Life, 2016 Annual Checklist. Disponível em: http://www.catalogueoflife.org/col/info/ac
Cavanaugh DP, Sternberg RV (2004). Analysis of morphological groupings using ANOPA, a pattern recognition and multivariate statistical method: A case study involving centrarchid fishes. Journal of Biological Systems. 12(2):137–67.
Cavanaugh DP, Wood TC (2002). A Baraminological Analysis of the Tribe Heliantheaesensulato (Asteraceae) Using Analysis of Pattern (ANOPA). Occas. Papers of the BSG. 17(1):1-11.
Chadwick A, Spencer L (2006). Turner Preliminary depositional model for an Upper Cretaceous Edmontosaurusbonebed. Journal of Vertebrate paleontology. 26:49A.
Crabtree GR (2013a). Our fragile intellect. Part I. Trends in Genetics. 29(1):1-3.
Crabtree GR (2013b). Our fragile intellect. Part II. Trends in Genetics. 29(1):3-5.
Crow JF (1997). The high spontaneous mutation rate: is it a health risk? Proc Natl AcadSci U S A. 94(16):8380-6.
Darwin C (1861). On the Origin of Species. 3. Ed. London: Murray. capítulo 14.
de Paula MO (2009). Podem os criacionistas aceitar a origem de novas espécies? Earth History Research Center. Disponível em: http://origins.swau.edu/papers/evol/marcia1/defaultp.html
de Queiroz K (2005). Ernst Mayr and the modern concept of species. Proc Natl AcadSci U S A. 102 Suppl 1:6600-7.
Dolgin E (2009). Human mutation rate revealed. Nature News (27 Ago. 2009). Disponívelem:http://www.nature.com/news/2009/090827/full/news.2009.864.html
Eakin CM (2014). Lamarck was partially right—and that is good for corals. Science. 344(6186):798-9.
Eldredge N, Stanley SM (1984). Living Fossils: Introduction to the Casebook. New York/Berlin: Springer-Verlag pp. 266-270.
Eldredge N (1986). Progress in Evolution? New Scientist. 110:54-57.
Eyre-Walker A, Keightley PD (1999). High genomic deleterious mutation rates in hominids. Nature. 397(6717):344-7.
Folha de S. Paulo (2006). Pássaros exibem seleção natural em tempo real. Folha de São Paulo (14/07/2006). Disponível em:http://www1.folha.uol.com.br/folha/ciencia/ult306u14848.shtml
Fondon JW, Garner HR (2004). Molecular origins of rapid and continuous morphological evolution. Proc Natl AcadSci U S A. 101(52):18058-63.
Foucaud (1897). Un Spartinainédit. Ann SocSci Nat Char Inf. 32:220–222.
Frair W (2000). Baraminology: Classification of Created Organisms. CRSQ Quarterly. 37(2):82-91.
Freeman S, Herron JC (2009). Análise evolutiva. 4. ed. orto Alegre: Artmed. 848p.
Fu W, et al (2013). Analysis of 6,515 exomes reveals the recent origin of most human protein-coding variants. Nature. 493(7431):216-20.
Gibson LJ (2002). A similaridade biológica e a teoria da evolução. In: 4º Encontro Nacional de Criacionistas, 24 a 28 de janeiro de 2002, São Paulo. Anais... São Paulo: Centro Universitário Adventista, campus São Paulo.
Gould SJ (1981). Evolution as Fact and Theory, Discover magazine 1981; 2(5):34-37; Reimpresso com permissãoem: Gould SJ. Hen's Teeth and Horse's Toes. New York: W. W. Norton & Company, 1994, pp. 253-262.
Grant PR, Grant BR (2006). Evolution of Character Displacement in Darwin's Finches. Science. 313(5784):224-6.
Groves H, Groves J (1880). Spartinatownsendiinobis. Rep Bot SocExch Club Br Id. 1:37.
Halliburton R, Gall GAE (1981). Disruptive selection and assortative mating in Triboliumcastaneum. Evolution. 35:829-843.
Herrel A, et al (2008). Rapid large-scale evolutionary divergence in morphology and performance associated with exploitation of a different dietary resource. Proc Natl AcadSci U S A. 105(12):4792-5.
Horner JR, Goodwin MB (2009). Extreme Cranial Ontogeny in the Upper Cretaceous Dinosaur Pachycephalosaurus. PLoS One. 4(10):e7626.
Hoskin CJ1, Higgie M, McDonald KR, Moritz C (2005). Reinforcement drives rapid allopatric speciation. Nature. 437(7063):1353-6.
Huey RB, et al (2000). Rapid Evolution of a Geographic Cline in Size in an Introduced Fly. Science. 287(5451):308-309.
Jerlström P (2000). Is the evolutionary tree turning into a creationist orchard? Journal of Creation. 14(2):11–13.
Johnson MW (1953). The copepod Cyclops dimorphus Kiefer from the Salton Sea. American Midland Naturalist. 49:188-192.
Junker R, Scherer S (1996). Evollução: um livro texto crítico. 4. Ed. Brasília, DF: Sociedade Criacionista Brasileira.
Kahle H (1999). Evolution: IrrwegmodernerNaturwissenschaft? 4. Ed. Bielefeld: Mindt, Edeltraud. 200p.
Keightley PD (2012). Rates and Fitness Consequences of New Mutations in Humans. Genetics. 190(2):295-304.
Kemp TS (1985). A Fresh Look at the Fossil Record. New Scientist. 108(1485):66-67.
Levinton JS, Chris MS (1980). A Critique of the Punctuated Equilibria Model and Implications for the Detection of Speciation in the Fossil Record. Syst Zool. 29(2):130–142.
Lightner JK (2016). Variation after the Flood. Creation Matters. 21(2):6-7.
Locey KJ, Lennon JT (2016). Scaling laws predict global microbial diversity. Proc Natl AcadSci U S A. 113(21):5970-5.
Lopes RJ (2015). Afobação faz cientistas classificarem fósseis de símios como hominídeos. Folha de S. Paulo. Disponível em: http://m.folha.uol.com.br/ciencia/2015/09/1676679-no-afa-cientistas-tem-classificado-fosseis-de-simios-como-hominideos.shtml?mobile
Lynch M (2010). Rate, molecular spectrum, and consequences of human mutation.Proc Natl AcadSci U S A. 107(3):961-8.
Marchant CJ (1963). Corrected chromosome numbers for Spartina x townsendii and its parent species. Nature. 199:929.
Marsh FL (1941). Fundamental Biology. Lincoln, NE: Published by the Author.
Marsh FL (1978). Variation And Fixity Among Living Things. A New Biological Principle.CRS Quarterly. 15(2):115-118.
Martens K (1997). Speciation in ancient lakes. Trends EcolEvol. 12(5):177-82.
Mora C, Tittensor DP, Adl S, Simpson AG, Worm B (2011). How many species are there on Earth and in the ocean? PLoS Biol. 9(8):e1001127.
Morrell V (1997). Catching Lizards in the Act of Adapting. Science. 276(5313):682-683.
Nachman MW, Crowell SL (2000). Estimate of the Mutation Rate per Nucleotide in Humans. Genetics. 156(1):297-304.
Nagalingum NS, et al (2011). Recent Synchronous Radiation of a Living Fossil. Science. 334 (6057): 796-799.
Prüfer K, et al (2013). The complete genome sequence of a Neanderthal from the Altai Mountains. Nature. 505:43-49.
ReMine WJ (1993). The Biotic Message. St Paul, MN: St Paul Science.
Ridley M (2006). Evolução. 3. ed. Porto Alegre: Artmed. 752p.
Robinson DA, Cavanaugh DP (1998). A quantitative approach to baraminology with examples from the primates. CRSQ Quarterly. 34:196–208.
Robinson DA (1997). A mitochondrial DNA analysis of the testudineapobaramin. Creation Research Society Quarterly. 33:262–72.
Romer AS (1966). Vertebrate Paleontology. 3rd Ed. Chicago: University of Chicago Press, pp. 347–396. In: Denton M. Evolution: A Theory in Crisis. Bethesda: Adler & Adler, 1986, p. 202.
Sadava D, Heller HC, Orians GH, Purves WK, Hillis DM (2009). Vida: A Ciência da Biologia. 8. Ed., v. 2. Porto Alegre: EditoraArtmed.
Sanford JC (2014). Genetic Entropy. 4. ed. New York: FMS Publications.
Scherer S (1993). Typen des Lebens. Berlin: Pascal.
Schwartz JH, Tattersall I (2015). Defining the genus Homo. Science. 349(6251):931-2.
Senter P (2010). Using creation science to demonstrate evolution: application of a creationist method for visualizing gaps in the fossil record to a phylogenetic study of coelurosaurian dinosaurs. Journal of Evolutionary Biology. 23:1732–1743.
Shabalina SA, Ogurtsov AY, Kondrashov VA, Kondrashov AS (2001). Selective constraint in intergenic regions of human and mouse genomes. Trends Genet. 17(7):373-6.
Shikano S, Luckinbill LS, Kurihara Y (1990). Changes of traits in a bacterial population associated with protozoal predation. Microbial Ecology. 20:75-84.
Snelling A (1997). Sedimentation experiments: Nature finally catches up! Journal of Creation. 11(2):125-126.
Teresi D (2011). Discover Interview: Lynn Margulis Says She's Not Controversial, She's Right. Discover Magazine, p. 68.
Trivedi BP (2000). Island mice may evolve faster: From one species to six. Genome News Network. Disponível em: http://www.genomenewsnetwork.org/articles/04_00/island_mice.shtml
Van Bocxlaer B, Hunt G (2013). Morphological stasis in an ongoing gastropod radiation from Lake Malawi. PNAS. 110(34):13892-13897.
Voje KL (2016). Tempo does not correlate with mode in the fossil record. Evolution. 70(12):2678-2689.
Weyrich A, et al (2016). Paternal intergenerational epigenetic response to heat exposure in male Wild guinea pigs. Mol Ecol. 25:1729-1740.
Whitmore J (2013a). Temporal Patterns in 'Living Fossils'. In: 4th Annual Research and Scholarship Symposium. Cedarville, OH. Anais... Cedarville, OH: Cedarville University. Disponível em: http://digitalcommons.cedarville.edu/research_scholarship_symposium/2013/poster_presentations/22/
Whitmore J (2013b). What about living fossils? Capítulo 12, pp.143-150. In: Ham K (Ed.). The New Answers Book, v. 4. Green Forest: Master Books.
Williamson PG (1981). Morphological Stasis and Developmental Constraint: Real Problems for Neo-Darwinism. Nature. 294:214-2015.
Wilmer JW, et al (2011). Catastrophic Floods May Pave the Way for Increased Genetic Diversity in Endemic Artesian Spring Snail Populations. PLoS One. 6(12):e28645.
Wilson G (2010). Classic Multidimensional Scaling isn’t the Sine Qua Non of Baraminology. Answers in Genesis. Disponível em: https://answersingenesis.org/creation-science/baraminology/classic-multidimensional-scaling-and-baraminology/
Wise KP (1990). Baraminology: a young-earth creation biosystematic method. In: Walsh RE (Ed.). Proceedings of the Second International Conference on Creationism. Volume II, Technical Symposium, pp. 345–360. Creation Science Fellowship.
Wise KP (1992). Practical baraminology. Creation ex Nihilo Technical Journal. 6(2): 122–37.
Wood TC, Cavanaugh DP (2001). A baraminological analysis of subtribe Flaveriinae (Asteraceae: Helenieae) and the origin of biological complexity. Origins. 52:7-27.
Wood TC, Wise KP, Sanders R, Doran N (2003). A Refined Baramin Concept. Occas. Papers of the BSG. 3:1-14.
Wood TC (2001). BDIST software, v. 1.0. Center for Origins Research and Education, Bryan College. Distributed by the author.
Wood TC (2002). A baraminology tutorial with examples from the grasses (Poaceae). Journal of Creation. 16(1):15–25.
Wood TC (2005). Visualizing baraminic distances using classical multidimensional scaling. Origins. 57:9-29.
Wood TC (2010). Baraminological Analysis Places Homo habilis, Homo rudolfensis, and Australopithecus sediba in the Human Holobaramin. Answers Research Journal 3:71–90.
Wood TC (2011). Using creation science to demonstrate evolution? Senter’s strategy revisited. Journal of Evolutionary Biology. 24(4):914–918.
Woodmorappe J (2000). The Karoo vertebrate nonproblem: 800 billion fossils or not. CEN Technical Journal. 14(2):47-49.
Woodruff DS (1980). Evolution: The Paleobiological View. Science. 208(4445):716-717.
Zimmerman EC (1960). Possible evidence of rapid evolution in Hawaiian moths. Evolution. 14:137-138.
Resumo - Estudos em genética de população conjugados com estatísticas genealógicas, revelam no recuo do tempo, um maior patrimônio genético nas espécies mães ancestrais das quais fomos derivados , bem como menor carga genética (entende-se por carga o acúmulo de mutações deletérias). Estes dois fatores verificados são justamente os que sustentam a hipótese de sobrevivência das especies ancestrais ao stress homozigótico existente nas nossas pequenas populações ancestrais,fato verificado nas etnias de biotipos semelhantes entre si e em estudos genéticos em desinovans (Nature, 2013). Destacamos que etnias habitando vastas áreas continentais em cima de fósseis de halogrupos dessemelhantes , sem estarem miscigenados com os mesmos, revela uma grande catástrofe global que os destruiu (fósseis caucasianos na china, fósseis negroides nas américas). A Incompatibilidade numérica taxonômica no registro fóssil em 543 milhões de anos , quando contrastamos este numero revelado em pobres arbustos filogenéticos fosseis, com o fato da especiação em tempo real, o grande numero taxonômico de especies e ricos arbustos filogenéticos, na nossa biodiversidade atual , nos levando a perceber que não houve tempo para muitas especiações/evoluções nos 543 milhões de anos, fazendo-nos verificar por esta perspectiva evolutiva a inexistencia deste tempo. Bem como há muita compatibilidade com o modelo criacionista biblico de que houve a extinção da maioria dos tipos basicos dos quais descendem todas as biodiversificações/especiações e adaptações fenopticas e genópticas atuais. Desta forma, compreendemos que a formação geológica do cambriano ao pleistoceno foi rápida devido a diversos fatores como 1. Pouco desgaste de trilhões de rochas erodidas em atrito com águas 2. Dimensão, extensão, espessura e largura das camadas geológicas contrastadas com menores extensões, larguras e espessuras conjugada a largura, das camadas geologicas formadas a partir do pleistoceno 3. Completude de grandes animais fossilizados e bem preservados com inexistencia de girafas, elefantes, hipopótamos e rinocerontes completos sendo fossilizados atualmente revelam eventos contextuais formadores do fanerozoico até o pleistoceno distintos e maiores que os atuais 4. Milhões de erosões muito violentas e energéticas, transportes e deposições, verificadas em todo o globo, revelam conjugação de eventos catastróficos globais, num tipo efeito-dominó, e defesa destas com a presença de grandes astroblemas como causas naturais das mesmas. 5. O acúmulo de tipos de sedimentos em determinadas camadas, bem como diversas linhas em plano paralelo das mesmas, revelam que houve separação hidrodinâmica automática pela diferença de composição fisico-química, assim como bem expressa a lei da segregação e extratificação espontanea (SEE) defendida por Marske e Berthaud.
|
|
3 TESES QUE DESENVOLVEMOS - QUALQUER CONTRIBUIÇÃO, CRITICA INTELIGENTE E REFUTAÇÃO, SEJAM BEM-VINDAS :
1) Paleontologia do T - Resumo
A hipótese de trabalho pode se resumir nesta pergunta: Quê diversidade e variações morfológicas se esperaria encontrar no registro fóssil a luz do comportamento bio-modificacional rápido e diversificador, escancarado na biodiversidade atual e até demonstrável nas especiações em tempo real?
Se encontramos trilhões de toneladas de especies nas camadas, não era de se esperar, caso fosse a fossilização um evento raro, uma maior diversidade dos achados, dado o fato de que especiação ocorre em tempo real e os arbustos filogenéticos da atualidade são tão ricos morfologicamente ? (já que o critério morfológico é o mais destacado na aferição taxonômica fóssil) não seria mais óbvio esperar distintos e não iguais ("especie em estase" como apontam os estudos paleontológicos pontualistas de Gould, Elredge e muitos outros grandes estudos da paleontologia?
A Incompatibilidade numérica taxonômica no registro fóssil em 543 milhões de anos, que é verificada quando contrastamos este pobre numero (300 mil espécies sendo ainda 84% artrópodes, revelado em pouco diversificados, e pobres arbustos filogenéticos) , com a riqueza diversificada causada pelo comportamento bio-diversificador evolutivo/adaptativo/derivado( verificado com especiações em tempo real).Este imenso contraste nos leva a duvidar da coerência da historia evolutiva.
O grande numero taxonômico de especies e diversificados arbustos filogenéticos, na nossa biodiversidade atual, nos leva a perceber que não houve tempo para muitas especiações/evoluções/adaptações no registro fóssil de supostos 543 milhões de anos, fazendo-nos constatar, por esta perspectiva evolutiva, a quase total inexistência deste tempo. Curiosamente a própria evolução contesta a historia da evolução.
Paralelamente podemos ver que há muita compatibilidade de falta de diversidade nos arbustos filogenéticos do registro fóssil, com o modelo criacionista bíblico que defende a extinção da maioria dos tipos básicos ancestrais dos quais descendem e derivam todas as biodiversificações/especiações e adaptações fenópticas e genópticas atuais.
2) Hipervariabilidade Ancestral - Resumo
Estudos em genética de população conjugados com estatísticas genealógicas, revelam no recuo do tempo, um maior patrimônio genético nas espécies-mães ancestrais das quais fomos derivados , bem como menor carga genética (entende-se por carga o acúmulo de mutações deletérias). Estes dois fatores verificados são justamente os que sustentam a hipótese de sobrevivência das especies ancestrais ao stress homozigótico (aumento de frequência de alelos deletérios quando há consagüinidade nos cruzamentos ) existente nas nossas pequenas populações ancestrais. Caso as especies ancestrais tivessem muitos alelos deleterios para compartilhar e pouco patrimônio diversificador e variabilizador, ela não sobreviveria ao stress homozigótico e nem derivaria as variações atuais http:// sodregoncalves.no.comunidades.n et/index.php?pagina=1399195911
3) Pouco tempo para formação geológica entre o Cambriano e o Pleistoceno - Resumo
A formação geológica do cambriano ao pleistoceno foi rápida devido a diversos fatores como:
1. Pouco desgaste de trilhões de rochas erodidas em atrito com águas em trilhões de demonstrações em cachoeiras, assoalho de rios, encostas dos mares, etc.
2. Dimensão, extensão, espessura e largura das camadas geológicas contrastadas com menores extensões, larguras e espessuras conjugada a largura, das camadas geologicas formadas a partir do pleistoceno
3. Completude de grandes animais fossilizados e bem preservados com inexistência de girafas, elefantes, hipopótamos e rinocerontes completos sendo fossilizados atualmente revelam eventos contextuais formadores do fanerozoico até o pleistoceno distintos e maiores que os atuais
4. Milhões de erosões muito violentas e energéticas, transportes e deposições, verificadas em todo o globo, revelam conjugação de eventos catastróficos globais, num tipo efeito-dominó, e defesa destas com a presença de grandes astroblemas como causas naturais das mesmas.
5. O acúmulo de tipos de sedimentos em determinadas camadas, bem como diversas linhas em plano paralelo das mesmas, revelam que houve separação hidrodinâmica automática pela diferença de composição fisico-química, assim como bem expressa a lei da segregação e extratificação espontanea (SEE) defendida por Marske e Berthaud. http:// igrejaadventista.no.comunidades .net/ index.php?pagina=1413735657 (*Esta tese é fruto de um mega debate de 9000 réplicas e tréplicas onde o autor foi a interface entre 4 professores de geologia (USP, UNISSINOS, UNB, UEB) e o mesmo ajudado por um ex-professor de geologia da USP).
6. Pedregulhos pequenos, arenosos, espalhados na superficie de imensos terrenos
7. Média organizacional de fósseis coincidindo com terrenos mais baixos a terrenos mais altos revelando sedimentação média primeira e posterior, segundo localização geográfica do habitat dos animais. Lembrando que imensos turbiditos formados em taludes, arrastariam seres mais rasos a SEE dos que forem transportados.
**Todas estas observações vieram de debates virtuais e foram extremamente testadas com pares especialistas nas áreas propostas.
|
Antes de ler o artigo necessario é que o leitor entenda deriva genética como provavelmente o motor mais predominante das modificações nas descendencias das especies, onde a seleção natural atuaria de forma menos determinante. Para tanto, colocamos aqui um artigo que esclarece este ponto. Boa leitura!
Deriva genética
Origem: Wikipédia, a enciclopédia livre.
Deriva genética, deriva gênica, deriva alélica,derivação genética ou ainda oscilação genéticaé um mecanismo microevolutivo que modifica aleatoriamente as frequências alélicas ao longo do tempo [1]. A deriva genética é um processo estocástico, não é possível prever a direção da mudança na frequência de um alelo causada pela deriva. Esse mecanimo resulta em perda de variação genética e na fixação de alelos em diferentes loci. Os alelos fixados pela deriva podem ser neutros, deletérios ou vantajosos. Nesses dois últimos casos, a trajetória da frequência alélica ao longo do tempo será determinada pela interação entre a deriva e a seleção natural.[2] O efeito da deriva é maior quanto menor o tamanho da população, podendo aparecer em diferentes momentos da história evolutiva da população:
• Em seu início (efeito fundador);
• Em um ou mais momento da sua história (efeito de gargalo);
• Em intervalos regulares (variação demográfica);
• Constantemente (há populações, em que a quantidade de indivíduos se mantém pequena durante anos).
Além da observação dos efeitos em populações pequenas, estudos demonstraram que em nível molecular ocorre um grande efeito da deriva, pois geralmente mutações neste nível dos organismos são neutras, e desta maneira não há efeito da seleção natural. Essa evidência é um dos pontos discutidos quando se compara o Neutralismo vs. Selecionismo, a cerca das mutações neutras, pois o único modo destas se fixarem é por deriva genética, deste modo isso é um dos pontos que se discutem quando estudiosos avaliam a importância de cada um desses eventos.

Representação esquemática da Deriva Genética
Deriva genética é a mudança na frequência dos alelos resultante de efeitos amostrais em populações finitas. Dentre os mecanismos evolutivos, aseleção natural é bem conhecida e popularizada, enquanto que a deriva muitas vezes pode causar certa dúvida. Como disse Suzuki et al. op. cit.: “O processo da deriva genética deveria parecer familiar, a qual é, na verdade, uma outra maneira de analisar o efeito do entrecruzamento em pequenas populações[...] Mesmo considerando-se como um entrecruzamento ou como uma amostra aleatória de genes, o efeito é sempre o mesmo. Populações não reproduzem exatamente as suas constituições genéticas; há um componente aleatório na mudança da frequência genética”.[3]
O conceito de Deriva genética foi tratado inicialmente, em 1929, por Sewall Wright, um dos fundadores do campo de genética de populações. Esses estudos iniciais sobre genética de populações estavam sendo agregados à teoria de Seleção Natural, e Wrigth analisava esses conceitos em pequenas populações, testando endogamia e observando a mudança de picos adaptativos, devida a deriva genética. Toda essa idéia de Wrigth não era aceita por Ronald Fisher, o qual dizia que a deriva genética tem um papel mínino na evolução, deste modo não afetaria os picos adaptativos.Kimura em 1968, com a Teoria neutralista da evolução, retomou o debate sobre a contribuição da deriva para explicar a variação genética encontrada em populações reais.[1]
[editar]Frequências alélicas
Os alelos sofrem deriva por períodos limitados. Eventualmente, a frequência dos alelos aumenta ou diminui de tal maneira que, ou atingem a frequência de 1 (único alelo representado na população), ou atingem a frequência nula (o alelo desaparece da população). Estes fenômenos são conhecidos como fixação e extinção de um alelo, respectivamente. No caso de a frequência de um alelo atingir o valor de 1, somente através de mutação esta poderá ser alterada novamente, desde que a população permaneça isolada,mas outro alelo poderá reaparecer caso ocorra fluxo gênico de outras populações[4]. A frequência de um alelo pode também ser modificada através da migração, onde novos indivíduos inserem variação alélica na população. Além da deriva genética, as mutações, migrações e a seleção afetam o equilíbrio gênico.
A permanência de um alelo numa população é governada pelo tamanho efetivo desta. Em populações de tamanho reduzido, poucas gerações podem ser suficientes até que um alelo sofra fixação por efeito da deriva. Em populações maiores, este efeito demora mais. Em média, um alelo neutro será fixado em  gerações, onde gerações, onde 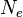 é o tamanho efetivo da população. é o tamanho efetivo da população.

Gráfico deriva genética aleatória
[editar]Tamanho populacional
É comum pensar que o tamanho populacional compreende todos os indivíduos que compõem determinada população (tamanho efetivo real, representado pela letra N), mas isso não é correto. O tamanho populacional efetivo (representado em fórmulas matemáticas por  ), são todos os indivíduos que se reproduzem e conseguem deixar descendentes, desta maneira, transmitindo os genes para frente (mantendo o fluxo gênico) e consequentemente mantendo a diversidade genética. Existem características para o tamanho populacional efetivo, as quais são: ), são todos os indivíduos que se reproduzem e conseguem deixar descendentes, desta maneira, transmitindo os genes para frente (mantendo o fluxo gênico) e consequentemente mantendo a diversidade genética. Existem características para o tamanho populacional efetivo, as quais são:
- Não há sobreposição de gerações;
- O número de indivíduos intercruzantes é o mesmo em todas as gerações;
- Todos os indivíduos são potencialmente intercruzantes;
- A união dos gâmetas é aleatória;
- Não há seleção, mutação ou ou migração;
- A média de descendentes, por indivíduo adulto, é de um individuo.
Desta maneira, uma população grande (valor alto de N) pode sofrer os efeitos da deriva genética, pois não são todos os indivíduos que se reproduzem efetivamente. O tamanho efetivo populacional pode ser calculado com diferentes fórmulas matemáticas as quais, dependem da estimativa da diversidade genética populacional. Mas a fórmula mais simples para estimar o tamanho efetivo é demonstrada a seguir, e ela considera a razão sexual entre adultos (maduros sexualmente), segue abaixo a equação[4]
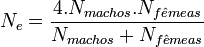
Onde:
- é o número total de machos na determinada população;

 é o número total de fêmeas na determinada população; é o número total de fêmeas na determinada população;- é o tamanho efetivo populacional.
O tamanho efetivo populacional é geralmente menor que a quantidade de indivíduos adultos, por exemplo, em uma população de 120 indivíduos, onde 100 são machos e 20 são fêmeas, o tamanho efetivo será de aproximadamente 67 indivíduos, caindo consideravelmente o tamanho populacional e desta maneira, há uma maior fragilidade aos efeitos da deriva genética, pois esta estará atuando sobre os 67 indivíduos e não sobre os 120. Geralmente diferenças entre o tamanho real e o tamanho efetivo são devido a:
- Espécies com razão sexual desigual (formação de haréns, ou até mesmo o exemplo acima citado);
- Com alta variação no tamanho da família ( diferença na quantidade de filhotes);
- Com número variável de indivíduos entre as diferentes gerações, o que também pode ser chamado de demografia oscilante.
| Proporção sexual |
Número de machos |
Número de fêmeas |
Ne |
| 1,00 |
50 |
50 |
100 |
| 3,00 |
75 |
25 |
75 |
| 0,33 |
25 |
75 |
75 |
| 9,00 |
90 |
10 |
36 |
| 0,10 |
10 |
90 |
36 |
| 99,00 |
99 |
1 |
3,36 |
| 0,01 |
1 |
99 |
3,36 |
| 5 |
100 |
20 |
67 |
[editar]Deriva Genética e Equilibrio de Hardy-Weinberg
O Equilíbrio de Hardy-Weinberg tem o pressuposto que determinada população irá se manter em equilíbrio gênico, ou seja, sem alterações na frequência gênica. Deste modo é possível a analise da porcentagem de indivíduos heterozigotos e homozigotos (A deriva genética tende a formar uma população homozigótica devido ao endocruzamento[4]). A deriva genética, por ser aleatória, desestabiliza populações que poderiam entrar no equilíbrio de Hardy-Weinberg, uma vez que altera uma regra para este acontecer, pois o mesmo diz que a população deve manter um constância, e a deriva faz com que a população desobedeça a regra e fuja do equilíbrio. Todavia, se a população for totalmente composta por indivíduos homozigóticos (devido a Deriva) ou está caminhando para essa característica, será possível a ocorrência do Equilíbrio de Hardy-Weinberg. Mas é importante lembrar que para populações reais, estar de acordo com o equilíbrio é quase improvável, devido as premissas que o mesmo necessita para ocorrer.[5]
[editar]Deriva genética e selecção natural
A deriva genética e a Seleção natural raramente ocorrem independentemente em populações naturais; estes dois fenómenos estão sempre a actuar numa população. No entanto, o grau em que cada alelo é afectado por estes dois fenómenos pode variar em função das circunstâncias.Nos dois processos é claro que há um efeito na evolução das populações, como dito anteriormente, entretanto sabemos que a deriva aleatória segue um caminho aleatório, enquanto que a seleção natural segue em um processo direcionado, além do mais a seleção natural permite uma adaptção dos organismos, deste modo os mais adaptados sobrevivem, pois a seleção busca eliminar os genes deletérios ou desfavoráveis, além de ignorar os genes neutros. Enquanto que a deriva, por ser aleatória, não há distinção entre genes bons e ruins.
Numa população com um tamanho efectivo elevado, a deriva ocorre muito lentamente, e a selecção actuante sobre um alelo pode de uma maneira relativamente rápida, aumentar ou diminuir a sua frequência (dependendo da viabilidade do alelo). Numa população com um tamanho efectivo reduzido, o efeito da deriva genética predomina. Neste caso, o efeito da selecção natural é menos vísivel pois o efeito da deriva muitas vezes se sobrepõe.
Desta maneira, não podemos dizer que apenas a seleção natural é um mecanismo evolutivo, e muito menos o único. Pois é evidente a ocorrência de deriva genética nas populações, sendo isso abordado em vários livros de genética e evolução [6], por exemplo: “Em qualquer população, algumas proporções de loci são fixas a um desfavorável alelo selecionado, porque a intensidade da seleção é insuficiente para sobrepor-se à deriva aleatória para a sua respectiva fixação. Um enorme ceticismo deveria ser mantido em direção às ingênuas teorias acerca da evolução que assumem que uma população sempre, ou quase sempre, alcança uma otimizada constituição somente por seleção. A existência de múltiplos picos adaptativos e a fixação aleatória de menos genes aptos são características integrais do processo evolutivo. Não podemos confiar à seleção natural a produção do melhor de todos os possíveis mundos”.[7] E “um dos mais importantes e controversos assuntos em genética de populações é a respeito da relativa importância da deriva genética e da seleção natural na determinação da mudança evolucionária. A chave da questão em jogo é se a imensa variedade genética, a qual é observável nas populações de todas as espécies, não é resultado da sobrevivência e reprodução (ou seja, é neutro), caso em que a deriva será o principal determinante, ou se a maioria das substituições genéticas afetam o fitness, caso em que a seleção natural é a principal força direcionadora. Os argumentos acerca deste assunto têm sido intensos durante a metade do século passado e estão pouco próximos da resolução, embora muitos diriam que o caso da deriva tornou-se progressivamente mais forte. Deriva, por sua própria natureza, não pode ser demonstrada positivamente. Para fazê-lo, precisar-se-ia, necessariamente, mostrar que a seleção definitivamente não operou, o que é impossível. Não obstante, muitas evidências indiretas foram obtidas, as quais pretendem favorecer a posição da deriva. Primeiramente, e de diversos modos, sendo a maioria convincente, é a evidência molecular e bioquímica...” [8]
Para ficar mais claro a diferença entre a Deriva e a Seleção Natural atuando em uma determinada população, um exemplo bem simples, poderá ajudar: Imagine uma população de 100 pessoas, as quais terão que escalar uma montanha. Neste grupo de individuos apenas 40 dessas pessoas possuem equipamentos adequados para a escalada, deste modo a seleção natural irá atuar de forma que apenas os mais preparados cheguem ao topo da montanha, assim os outros 60 (sem equipamentos adequados, ou seja, menos adaptados) irão ser eliminados. Todavia, se ao momento em que todos os indivíduos estiverem escalando, pedras comecem a cair, deste modo tanto pessoas com equipamentos apropriados, tanto pessoas sem equipamentos serão atingidos, aleatoriamente, de forma que mesmo os participantes equipados sofram consequência e sejam eliminados não chegando ao topo. Deste modo houve ação da oscilação genética.
[editar]Deriva genética e as populações
A deriva pode ter efeitos dramáticos na história evolutiva das populações. São ditos como exemplos da deriva genética os eventos:
- Efeito Gargalo;
- Efeito Fundador;
Exemplos estes, explicados abaixo:
Quando ocorre o efeito de gargalo, a deriva genética pode resultar em rápidas e dramáticas alterações nas frequências alélicas. Este facto ocorre de maneira independente da selecção natural, a chances de sobrevivência dos indivíduos que o sofrem é totalmente aleatória também e não há vantagem genética atuando. Nestes casos, muitas adaptações benéficas poderão ser eliminadas da população. Pensa-se que o efeito de gargalo tenha ocorrido por diversas ocasiões na história da evolução humana. Um exemplo de população que sofreu efeito gargalo são os Elefantes Marinhos do Norte, os quais devido a caça predatória, a população foi reduzida a 30 indivíduos. E após 25 anos, chegou a 10.000 indivíduos.[4] Neste último momento, o qual ocorreu um aumento populacional, dizemos que ocorreu o efeito do fundador. Desastres ecológicos, como incêndios florestais, desmatamento, inundações, etc., podem fazer populações sofrerem efeito gargalo e até mesmo efeito fundador.
Similarmente, as populações migrantes, como por exemplo saída do continente em busca de uma ilha, poderão sofrer o chamado efeito do fundador. Este efeito é muitas vezes responsável por uma frequência elevada de algumas doenças genéticas. Podemos relacionar os dois efeitos: O efeito gargalo pode levar uma população ao efeito fundador, entretanto, o contrário não ocorre.
Em populações que estão em processo de extinção, a deriva genética pode ser drástica, elimando de uma vez a população a qual sofre o seu efeito. Além disso, já que em cada população podem ser diferentes os alelos que se perdem e se fixam, a deriva faz com que duas ou mais populações da mesma espécie tendem a se diferenciar geneticamente.
Um outro fator de grande influência para ocorrer os efeitos da deriva aleatória, é aEndogamia, que seria o cruzamento entre parentes, endocruzamento ou consanguinidade. O extremo da endogamia é a auto-fecundação, a qual geralmente é observada em espécies vegetais. A endogamia influência muito a história da população, pois a consequência deste evento é a redução reprodutivo, bem como a sobrevivência da prole (Depressão endogâmica). Devido a endogamia a quantidade de heterozigotos é reduzido, mas não muda a frequência de genes e concomitantemente há um aumento dos indivíduos homozigotos, deste modo a exposição e expressão de genes deletérios se torna mais fácil. A endogamia é inevitável em populações pequenas, pois com o passar do todo todos indivíduos se tornam aparentados.
[editar]Ligações externas
- - Seleção natural e deriva genéticadreyfus.ib.usp.br
- [1] - www.icb.ufmg.br
- [2] - Simulação gráfica da Deriva Genética
- [3] - Interação entre Deriva e Seleção Natural
- [4] - Teorias e Modelos em Genética de Populações: Um exemplo do uso do Método Axiomático em Biologia
- [5] - Genismo
Referências
- ↑ a bRidley, Mark. Evolução (3ª edição) Tradução: Henrique Ferreira, Luciane Passaglia, Rivor Fischer - Porto Alegre: Artmed, 2006 ISBN 978-85-363-0635-3
- ↑ FREEMAN, Scott; HERRON, Jon C. Análise Evolutiva (4ª edição) - Porto Alegre: Artmed, 2009. 831p.
- ↑ Suzuki et al. op. cit.
- ↑ a b c d Futuyma, Douglas. Biologia Evolutiva 2ª edição - Tradução: Mario de Vivo; Fabio de Melo Sene. Ribeirão Preto: Sociedade Brasileira de Genética/CNPq, 1992 ISBN 0-87893-183
- ↑ http://lineu.icb.usp.br/~bbeiguel/Genetica%20Populacoes/Cap.1.pdf
- ↑ Moran, L.. Deriva Genética Aleatória. Projeto Evoluindo - Biociência.org.
- ↑ Suzuki, D.T., Griffiths, A.J.F., Miller, J.H. and Lewontin, R.C.. An Introduction to Genetic Analysis 4th ed. W.H. Freeman, New York 1989
- ↑ Harrison, G.A., Tanner, J.M., Pilbeam, D.R. and Baker, P.T.. Human Biology 3rd ed.Oxford University Press 1988 pp 214-215
|
Qualquer professor de biologia sabe que pequenas populações não sobrevivem ao "stress homozigótico" (gerando cruzamentos cada vez mais próximos e consanguinos no recuo das populações genalogicamente ancestrais e/ou isoladas) , exceto raras excepções que até espelham o que vamos dizer aqui... todos nós precisamos confirmar nas estatisitcas que até nós humanos ( uma das especies mais aptas do planeta) éramos bem pouco numerosos a 3, 4, 5 mil anos atrás..os crentes em darwinismo historico imploram por uma população inexistente de +-10 mil pra poder sobreviver ao stress homozigotico consanguino no inicio ...mas isso é pedir uma hipótese cada vez mais improvável quanto mais voltarmos a MÍSEROS 4 mil anos atrás...ou seja, o que queremos dizer é que as populações ancestrais precisariam, pra sobreviver ao stress homozigótico e consanguinidade acentuada nos cruzamentos, quando pequenas (e isso vale para diversas pequenas familias isoladas neste vasto planeta), ter um super DNA para sobreviver, a exemplo das raras especies que conseguem tal façanha ainda hoje...UM SUPER DNA seria poucos ALELOS DELETERIOS para compartilhar E um DNA RICO EM VARIABILIDADE. Isso prova que Bíblia iniciando com um casal perfeito criado pelas mãos de Deus, sem defeitos, sem alelos deleterios para compartilhar e necessaariamente super variaveis é a UNICA RESPOSTA , O UNICO CAMINHO E A UNICA VERDADE. mas a Biblia ainda brinca de prever todas as observações corretas do darwinismo no campo da biologia funcional e não biologia extrapolada a tempos idos... A Bíblia relata que feita a população inicial na terra , houve uma mortandade drástica e consequentemente uma queda brusca de variabilidade deixando apenas 4 casais descendentes(o que combina com 274 lendas do diluvio em TODAS as civilizaçãoes antigas do globo da terra com até 42% de média de semelhança na comparação destes relatos arqueologicos chineses, egipcios, etc) ..Amigos, a Bíblia sem saber nada de efeito gargalo e fundador de sub-especies, acaba por relatar essa ocorrencia ao descrever a queda vertiginosa de longevidade nas populações pos-diluvianas (veja Genesis 6 a 12) mostrando claramente que houve uma SUB-ESPECIAÇÃO na linhagem humana ...Pessoal, quem ainda ainda se discute criacionismo x evolucionismo...é porque não sofreu esta atualização..aperte a techa CRIACIONISMO UP TO DATE, leiam PELO MENOS um livro técnico do criacionismo (indico http://www.scb.org.br/livros/imagens/Evolucao.jpg ) e tudo se resolve!!!
 NETO, G B Sodré, Hipervariabilidade Ascendente. Monografia apresentada em exigência da materia de Metodologia Cientifica, do curso de pós graduação em Ciência da Religião, da Faculdade INTA . Maio de 2008
NETO, G B Sodré, Hipervariabilidade Ascendente. Monografia apresentada em exigência da materia de Metodologia Cientifica, do curso de pós graduação em Ciência da Religião, da Faculdade INTA . Maio de 2008
Se originou por meio de debates com especialistas na área em 2007, e vem sendo aprimorado, acrescentado, corrigido e melhorado desde então. Até meados de 2010 recebeu constantes réplicas e tréplicas de mais de 30 estudiosos em genetica, incluindo especialistas em estatistica populacional, biologia evolutiva, arqueologia, etc.. numa especie de banca virtual que durou vários anos. Depois de 2010 recebeu pouquissimas objeções e muitas fugas daqueles que defendiam o modelo "evolucionista historico" (Mayr chama de biologia historica a biologia evolutiva funcional + geocronologia).
Agradecemos a todos que de forma direta ou indireta contribuiram para o mesmo, e esperamos maiores oportunidades para poder melhorar , organizar, sistematizar , divulgar. Agradecemos a gestão do portal da educação que de forma livre, permitiu diversas discussões, representando assim o espirito universitario.
 "Variação do DNA humano se aproxima da linha temporal bíblica
"Variação do DNA humano se aproxima da linha temporal bíblica
Um estudo publicado na revista Science pode comprovar a verdade Bíblica do Dilúvio e da diversificação acelerada dos seres humanos depois desse episódio. Isso porque a pesquisa mostra que a variação do genoma humano começou há aproximadamente 5 mil anos atrás.
Os cientistas já sabiam que cada pessoa é distinta da outra, com exceção dos gêmeos idênticos, mas o que eles quiseram descobrir com essa nova pesquisa era quando essa distinção começou. Para isso foram analisadas as sequenciais do DNA de 15.585 regiões genéticas codificadas de proteínas do genoma humano.
Esse material genético foi retirado de 1.351 americanos com descendência europeia e 1.088 com descendência africana. Com esses dados em mãos a equipe pode então examinar o percurso da variação genética através do tempo, dando como resultado a seguinte frase: “O limite máximo provável para o crescimento acelerado foi há 5,115 anos atrás”.
Para quem defende a Terra tem milhões de anos fica a pergunta: Por que com tantos milhões de anos a diversificação genética dos humanos só passou a acontecer nos últimos 5.000 anos?
Enquanto os defensores da evolução das espécies tentam encontrar uma resposta, a pesquisa aproxima o fato com os relatos sobre a história de Noé que aconteceu há cerca de 4.500 anos, se ajustando ao tempo máximo que os pesquisadores estimam".
http://noticias.gospelprime.com.br/variacao-do-dna-humano-se-aproxima-da-linha-temporal-biblica/
Especiação para criacionismo vai alem dacapacidade de uma especie se tornar uma sub-especie para poder se adaptar a contingencias diversas.
E Mayr, um dos maiores cientistas da evolução, ao falar sobre as dificuldades da especiação assim expressa:
"Especiação é um processo perigoso. O empobrecimento do patrimônio gênico e a instabilidade genética que a acompanham, pode conduzir com maior probabilidade ao desastre do que ao sucesso. Apesar da maioria das espécies insipientes não se extinguir, uma espécie ocasional não apenas pode completar o processo, como pode ter sucesso na penetração de um nicho ou zona adaptativa novos." Ernest Mayr, populações, Espécie e Evolução p.353.
Se Mayr chama especiação de um processo perigoso, salta-nos idéia de tendencia média entrópica nas especiações. Portanto podemos inferir uma tendencia média de ordenação genética nas ascendencias, ascendentes com maior variabilidade e menor acumulo de carga genetica (mutações). Isso nos lembra o que já falámos em 2006, de especiações que deram certo em sentido apenas relativo as circunstancias e nã em sentido de qualidade da especie ou sub-especie. Medimos qualidade por alta taxas de variabilidade e baixos acumulos de mutações deleterias ou não (Carga).
O neodarwinismo , que é a maestra regente da cosmovisão da biologia convencional, não tem COMO DEMONSTRAR , exceto por calculos baseados em hipoteses "AD HOC" e formulas escolhidas, como as populações ancestrais conseguiram sobreviver ao que se esperaria nas populações ancestrais: Uma suposta homozigose cada vez mais acentuada quanto mais voltarmos na ascendencia (devido ao ÓBVIO afunilamento estatistico populacional). Isso se levarmos em conta um uniformismo em relação ao DNA, como se ele fosse muito semelhante ao atual e sofresse lentas taxas de mutação. Mas veremos neste trabalho que para que a população humana pudesse sobreviver necessario seria um DNA hipervariavel e com baixissima carga genetica. (mutações).
"Em ciência e filosofia, ad hoc significa a adição de hipótese(s) estranha(s) a uma teoria para salvá-la de ser falseada. Hipóteses ad hoc compensam anomalias não previstas pelas teorias em sua forma original, ainda não modificada. Cientistas são frequentemente céticos sobre teorias que as apresentam frequentemente, não suportando ajustes para sustentá-las. Hipóteses Ad hoc são frequentemente características de pseudocientistas.[1] Muito da produção científica reside sobre a modificação de hipóteses existentes ou teorias mas estas modificações são distintas de hipóteses ad hoc no que as anomalias são explicadas propondo um novo significado daquilo que é real e observável". http://pt.wikipedia.org/wiki/Ad_hoc
Logo, torna-se extremamente e indispensavelmente necessario a necessidade de hipervariabilidade e menor carga (acumulo de mutações) nas populações ancestrais para populações cada vez mais pequenas, para que mesmo com numeros populacionais baixos ou baixíssimos e insuportaveis pela perspectiva uniformista (como se o DNA fosse quase o mesmo ao atual) , conseguisse sobreviver a cruzamentos consaguinos e conseguisse permitir inumeras adaptações/variações assim sem o risco do stress homozigótico.
Os modelos iniciais da população humana e de quaisquer outros "tipos basicos" ou ramos de comprovada possibilidade de cruzamentos, são matematicamente IRREAIS, pois as populações ancestrais são cada vez menores e a biologia precisa, no caso de humanos, de pelo menos 10 mil individuos (ad hoc)..passou daí a Teoria de origem das especies quando falseada em seu inicio verifica-se simplesmente insuficiente e falsa. Não podemos de forma alguma simplesmente aceitar que começamos com 10 mil de alguma forma, quando a estatistica genealogica e dezenas de outros criterios nos falam exatamente o contrario, que éramos decrescentes em diversos pontos isolados e tambem em halogrupos.
A única saída que a genetica de populações possui é um modelo que contemple diferenciação genetica ascendente hipervariavel, com baixissima carga genetica, pois somente uma alta variabilidade e baixissimo fitness (não variabilidade adaptativa é claro..provocada por mutações, reações imunologicas, etc) , poderia SUPORTAR e sobreviver com população pequena em diversos pontos isolados onde se fundaram etnias.
Como este artigo originou de debates com especialistas na área em 2007, e vem sendo aprimorado, melhorado, corrigido, desde então, respondeu um doutorando especialista em genetica de populações o seguinte:
"Sim, a homozigose é favorecida em populações MUITO pequenas. Isso é fato. Porém, é fato para populações MUITO pequenas. Quando dizemos que uma determinada população pequena se isola de uma população mãe e daí o processo de especiação vê-se favorecido, não estamos querendo dizer que são populações abaixo de um nível demográfico crítico para a sua saúde genética. Nestes casos, essas populações simplesmente perecem, COMO A HISTÓRIA DA VIDA NA TERRA BEM DEMONSTRA".
Respondi que devemos lembrar que todas as populações ancestrais obrigatoriamente são cada vez mais pequenas, todas sem exceção - Todo inicio do inicio é bem pequeno, em diversas fases anteriores esta é a tendencia. Inclusive para se criar uma etnia com baixa variabilidade, é necessario populações muito pequenas, talvez poucos casais. Mas a TE precisa de limites em suas hipóteses para dar certo.
Phylogeny reconstruction is the process of inferring evolutionary relationships from molecular sequences, and methods that are expected to accurately reconstruct trees from sequences of reasonable length are highly desirable. To formalize this concept, the property of fast-convergence has been introduced to describe phylogeny reconstruction methods that, with high probability, recover the true tree from sequences that grow polynomially in the number of taxa n. While provably fast-converging methods have been developed, the neighbor-joining (NJ) algorithm of Saitou and Nei remains one of the most popular methods used in practice. This algorithm is known to converge for sequences that are exponential in n, but no lower bound for its convergence rate has been established. To address this theoretical question, we analyze the performance of the NJ algorithm on a type of phylogeny known as a 'caterpillar tree'. We find that, for sequences of polynomial length in the number of taxa n, the variability of the NJ criterion is sufficiently high that the algorithm is likely to fail even in the first step of the phylogeny reconstruction process, regardless of the degree of polynomial considered. This result demonstrates that, for general n-taxa trees, the exponential bound cannot be improved.
M. Lacey and J. Chang. A signal-to-noise analysis of phylogeny estimation by neighbor-joining: Insufficiency of polynomial length sequences. Mathematical Biosciences 199: 188-215, 2006
No caso de humanos, por diversos prismas, podemos perceber isso: Fiz uma lista de alguns poucos fatores que denominamos fatores de afunilamento populacional, que podem ser citados como fontes para uma hipótese de baixa população humana a pouco tempo atrás, são eles:
1.Línguas de afunilam a 3, 4 , 5 troncos básicos em 4-8 .000 AC com cada vez menor numero de palavras, de nomes, nomes cada vez mais iguais e cada palavra com cada vez maior numero de diferentes significados (homonimas)2.População humana por calculo estatístico inicia a 4-8.000 anos
3.Costumes e culturas cada vez mais semelhantes no recente passado
4.Genealogia (Yale, Nature aponta a possibilidade estattistica de apenas um ancestral comum aos vivos a "apenas" 3000 anos atrás )D. Rohde, S. Olson, and J. Chang. Modelling the recent common ancestry of all living humans. Nature, 431:562-566, 2004If a common ancestor of all living humans is defined as an individual who is a genealogical ancestor of all present-day people, the most recent common ancestor (MRCA) for a randomly mating population would have lived in the very recent past (ref). However, the random mating model ignores essential aspects of population substructure, such as the tendency of individuals to choose mates from the same social group, and the relative isolation of geographically separated groups.Here we show that recent common ancestors also emerge from two models incorporating substantial population substructure. One model, designed for simplicity and theoretical insight, yields explicit mathematical results through a probabilistic analysis. A more elaborate second model, designed to capture historical population dynamics in a more realistic way, is analysed computationally through Monte Carlo simulations. These analyses suggest that the genealogies of all living humans overlap in remarkable ways in the recent past. In particular, the MRCA of all present-day humans lived just a few thousand years ago in these models. Moreover, among all individuals living more than just a few thousand years earlier than the MRCA, each presentday human has exactly the same set of genealogical ancestors.
http://www.stat.yale.edu/~jtc5/papers.html
5.Historia inicia em 4-8 mil anos, hieróglafos que deram inicio a escrita datados neste inicio comum (4-8 mil)
6.Número de Cidades reveladas pelas escavações vai decaindo muito
7. Referencias ao ciclo semanal de 7 dias em diversas culturas ancestrais
8. Igualdades culturais e religiosas apontam para poucos em tempos arqueologicos
9. Costume claro em todos os povos antigos de casamentos consanguinos/primos com baixa incidencia de relatos de síndromes (o que indica baixo acumulo de mutações deleterias e menor frequencia de alelos deleterios em casamentos consanguinos)
10.Nomes iguais
11.Deuses iguais (as culturas mais antigas vão diminuindo o numero de deuses )
12.Geografia de inicio ancestral apontando para oriente medio e norte da africa
Mas a TE em centenas de casos onde fica implorando possibilidades..neste caso ela implora pelo menos uns 10000 individuos pra poder começar os calculos...e ela é super ajudada- um monte de trabalhos e calculos a favor desta IMENSA carencia...Exatamente por isso, denominamos a TE como uma forte e bem fundamentada alienação da realidade, que por agregar muitos estudos e fundamentações, acaba por torcer os dados a sua erronea cosmovisão e não deixar que os mesmos falem por si. O mesmo pode se acusar quanto ao criacionismo bíblico, porem, em termos de cosmovisão, o criacionismo não carece de muitos aspectos filosoficos que o ateismo materalista evolucionista possui, portanto, está , em termos de ajustes muito mais adequado a realidade do que o que o materialismo impõe.Um estudo do Zoologo holandez DUYVENE DE WIT observa que "quando uma população marginal abre caminho para um novo habitat , ela não pode levar consigo todos os genes da população materna, mas apenas uma parte.Cada nova raça ou especie que se origina de uma outra possui portanto, um pool genico mais pobre." se o processo de especiação se repetir vezes consecutivaqmente, surgirão por fim, especies cujos pools genicos estão tão completametne esgotados que bastam alterações insignificantes das condições ambientais para que elas venham a se extinguir"."esforços para se adaptar a modificações ambientais como resultado de possibilidades insuficientes de recombinação levram, por fim, a um estado genetico minimo. Se este limite minimo for ultyrapassado, não haverá mais possibilidade de sobrevivencia. Por esta razão, o destino tragico e irrevogavel das especies ou raças especializadas , extremamente adaptadas, é a MORTE GENETICA. "Citado por KAHLE -1984 em "evolution - irrweg modeme naturwissenschaft? Bielifeld".O biólogo Fabio ponderou sobre especiação alopátrica ao que respondi:Mesmo que na alopátrica ocorra um empobrecimento menor de pool genico, ainda assim vai ocorrer. Exceto se todos os alelos migrarem para a isolada, o que me parece ser IMPOSSIVELEle então respondeu exigindo novamente uma população grande, "Parece, mas não é. Basta uma população original grande o suficiente".mas vejam: exigir população grande na ancestralidade é uma falha cada vez mais acentuada, pois quanto mais voltamos no tempo, TODAS ou quase todas populações tendem a cair em numeroLÔNNING (1993) cita eurioquia relacionando antepassados com capacidade de povoar e se adaptar a varios ambientes.E descendentes (estenoquia) como podendo povoar , mas se adaptar a poucos habitats, devido ao aumento de frequencia de especializados.Esta tendencia vemos na natureza e nas extinções daqueles que não se adaptam mais.Em azul, respondi a tres contestações:"ocorrendo na natureza com a necessidade de hipervariabilidade e menor carga (acumulo de mutações) nas populações ancestrais para populações cada vez mais pequenas e mesmo assim sem terem o risco do stress homozigótico (Sodré)""vamos la´de novo com algumas perguntas:1) como houve a hipervariabilidade já que tu discarta as mutações?as muitas derivas genetica e seleções naturais que as populações sofreram no planeta ao longo do tempo, revelam que houve empobrecimento do pool genico, em muitas das diversas adaptações e especiações que as especies-mães sofreram. Os diversos efeitos gargalos e fundadores ocorridos diante de catastrofes foram acompanhados por empobrecimentos de pool genicos. Logo, calculamos diante de varias evidencias destes processos empobrecedores, que éramos quando especies-mães, mais ricos, mais hipervariaveis , mesmo em populações ancestrais pequenas.2) vemos populações aumentando, como se dá esse choque entre populações já que a tua proposta é que as populações diminuem?minha não, das estatisticas populacinais ascendentes e dos varios outros prismas que nos revelam que éramos muito poucos a poucos milhares de anos
http://www.portaleducacao.com.br/forum/forum_posts.asp?TID=6373) como se evita a homozigose já que a população é pequena?Há casos de alta variabilidade em populações pequenas, este casos raros lançam luz sobre a grande necessidade deste perfil nas populações primitivas
Quadro que ilustra diversos ramos gerando variações fenopticas e especiações cada vez mais afiladas (com menor patrimonio) e mais frágeis (aumento de carga genética negativa de alelos deleterios) culminando na extinção da especie. Excetua-se quadros onde especies primas se reencontram depois de segregaçãões pretéritas, onde ocorre o aumento de patrimonio e variabilidade, inclusive imunologica. Contudo, esta realidade de tendencia positiva é vista nestes contextos, em momentos da historia de uma população.
ADÃO E EVA E A GENÉTICA DE POPULAÇÕES
EVIDENCIAS DE LITERALIDADE EM TEMPO E QUANTIDADE , NO ESTUDO DE GENÉTICA DE POPULAÇÃO, ATRAVÉS DE ESTUDO DA VARIABILIDADE, CARGA NEGATIVA E PATRIMÔNIO GENÉTICO NAS POPULAÇÕES ANCESTRAIS
por
Sodré Gonçalves
1.Introdução
Cap 1 ..Genealogia e Ancestralidade Comum
1.2 Analisando Estudos sobre a Eva Mitocondrial (DNAmt)
Cap 2 -Baixa Variação Genética , doenças genéticas e Perigo de extinção
2.1 Os Perigos da Endogamia Continuada
2. 3 Situações Ideais Genéticas para Sobrevivência das Espécies Ancestrais
Cap 3 -Aumento de Potencial de Variabilidade nos genótipos ancestrais
3.1 Análise de Progressões Populacionais Convencionais
3. 2 Perímetro de convergência temporal
3.3 Análise de Parâmetros para Simulações de Sobrevivência de Espécies Ancestrais
3.4 Justificativas que Demonstram Longevidade nas Populações Ancestrais
Cap. 4. Conclusão
Resumo: O Estudo de Baixas populações demonstram risco de extinção por baixa variabilidade genética, onde os defeitos genéticos herdados aparecem com maior frequência. Esta realidade observada e testada, presente em inúmeros exemplos, nos leva a questionarmos o modelo que descreve uma média de complexificação das espécies no histórico fóssil. O inicio de populações de especies ancestrais necessita de uma realidade genética hipervariável e com um minimo de defeitos deleterios, e ao mesmo tempo, com a demonstração de vários casos onde especies-mães possuem menor carga genética de defeitos e maior patrimonio genético, nos faz observar que o código genético nas populações ancestrais fosse cada vez mais hipervariável, com menor defeito ou acúmulo de mutações deleterias e com maior patrimônio e complexidade genética, excetuando aspectos de imunidade, fruto de evolução de soluções geneticas de organismos selecionados frente as contingencias da vida. Em conformidade com isso, estudos demonstrando ancestralidade comum pelo DNAmt (DNA mitocondrial), pelo cromossomo Y e por cálculo estatístico, revelam individuo macho e fêmea como originador de toda humanidade vivendo não mais que poucos milhares de anos atrás, remetendo-nos invariávelmente ao que os escritos antigos já falavam de alguma forma.
Introdução
GENEALOGIA E ANCESTRALIDADE COMUM

Marco Antônio Zago e Wilson Silva Jr., da Faculdade de Medicina de Ribeirão Preto da Universidade de São Paulo (USP) escrevendo sobre a origem das populações de indios nas 3 américas, e através de estudos no DNAmt (DNA mitocondrial), deduziram, entre outras assertivas:
"É como se todos os haplogrupos tivessem aparecido mais ou menos ao mesmo tempo."
"Como os ameríndios apresentam menos diversidade genética do que os asiáticos, os pesquisadores acreditam que os nativos da América sejam o resultado de um processo evolutivo chamado de efeito gargalo: a partir de poucos indivíduos - leia-se baixa diversidade genética - origina-se uma população enorme que vai colonizar uma grande área - as três Américas, no caso".
http://www.revistapesquisa.fapesp.br/novo_site/extras/imprimir.php?id=1853&bid=1
Observamos que o afunilamento nos ascendentes não é uma observação isolada, mas se comunica multidisciplinarmente com varios raciocinios, testes e até mesmo centenas de fontes arqueológicas, e aponta não somente para baixas populações (com risco de homozigose continuada sempre aumentando quanto mais voltarmos), como também nos exige o desafio de respondermos a difícil pergunta de como aumentariam sua variabilidade para sobreviverem e justificar as centenas de variações e especiações atuais?
Para isso, algumas respostas se destacam como na matéria "Cientistas dos EUA têm nova teoria sobre evolução humana"
http://www.bbc.co.uk/portuguese/ciencia/021224_hominideobg.shtml
"Há sinais de que houve cruzamento entre espécies nesse período"
Mas....
"Esta nova pesquisa sugere que aconteceram alguns cruzamentos, mas, como os próprios autores reconhecem, esses cruzamentos podem ter ocorrido em dose limitada. O que realmente aconteceu continua sendo uma pergunta sem resposta", acrescentou o cientista".
Uma das maneiras de se estabelecer que houve cruzamentos entre supostos antepassados comuns ao homem e aos chipanzés é a localização de retrovirus, porem este raciocinio ignora que existe apenas parcialmente aleatoriedade na inserção dos mesmos pois " os elementos transponíveis se inserem apenas entre determinados pares de base (e isso depende do transposon, uns são mais específioos que outros), o que dita isso são as características físico-químicas das enzimas que os inserem nesses lugares"
http://www.orkut.com/CommMsgs.aspx?cmm=26688928&tid=2597695239025054347&na=4&nst=141&nid=26688928-2597695239025054347-5202024475795995748
Neste artigo, vemos que uns se inserem aleatoriamente e outros "preferem" certas sequencias.
Transposons are mobile genetic elements that insert themselves into genomic DNA at essentially random locations, although different classes of transposons can have preferences for certain insertion site sequences. For example, random mutagenesis of viral BACs can be accomplished using a donor plasmid (such as a Tn1721-derived insertion from a modified TnMax plasmid [42, 43, 76-78]) with a transposon that has a preference to be inserted into plasmid (rather than genomic) DNA [10]. (...)
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2965428/?tool=pubmed
Ou seja, nem sempre igualdades de retrovirus podem indicar parentesco e ancestralidade comum.
Estudos e testes efetuados pelo bioquímico Frank Sherer da alemanha, demonstram em testes feitos entre mais de 2000 especies e generos distintos , que para se estabelecer ancestralidade comum deveria haver ao menos um inicio de um embrião em cruzamentos artificiais, pelo menos uma coordenação cromossomica ajustavel. Ele procurou saber se isso ocorreu em tentativas artificiais entre homem e chipanzés, gorilas, etc... e não encontrou nem um inicio de formação embrionaria. "Evolução, Livro Texto Crítico"
Lembrando que existe certos setores do DNA onde acontecem mais erros de copias (por exemplo na heterocromatina) a polimerase do DNA comete mais erros. Ou seja, não reparam os erros outrora acontecidos, o que poderá coincidir mutações "justamente" em semelhantes locais, o que enfraquece a "coincidência" ainda mais. Certos setores concentram elevadas taxas de mutação também.
Ou seja, não apenas a pergunta sobre nossos ascendentes fica sem resposta como tambem os sinais de ascendência comun com chipanzés e o difícil cruzamentos entre especies primas , ficam mais fracos ainda, se não bastasse a fraqueza do proprio argumento levando em conta a dificuldade que é descendentes férteis, e a necessidade imensa de maior parentesco e semelhança com os simios, o que pesquisas recentes descartam cada vez mais esta possibilidade como vemos neste artigo da Science 29 June 2007: Vol. 316. no. 5833, p. 1836
Jon Cohen, EVOLUTIONARY BIOLOGY: Relative Differences: The Myth of 1%.
http://www.sciencemag.org/cgi/content/summary/316/5833/1836
Para verificar mais as numerosas diferenças genéticas, ver 23
http://www.orkut.com/CommMsgs.aspx?cmm=14251159&tid=2544078963181814212&kw=o+mito+do+1%25
Para quem quiser ver o impacto desta edição em comunidades defensoras da ancestralidade comum com chipanzés, acesse:http://www.orkut.com/CommMsgs.aspx?cmm=184959&tid=2552443873686910298&kw=o+mito+do+1%25
Especiação
http://www.colegiosaofrancisco.com.br/alfa/idade-do-gelo/formacao-de-novas-especies.phpOs autores evolucionistas insistem em apelar para mecanismos de especiação para justificar o surgimento da especie humana e de todos os seres vivosMas percebemos que estes mecanismos precisariam sempre de uma população anterior, como temos em http://www.simbiotica.org/especiacao.htm"Evolução divergente - ocorre quando duas populações se separam e acumulam diferenças que impossibilitam o cruzamento entre elas, originando novas espécies;#Radiação adaptativa - a partir de uma espécie inicial, verifica-se uma ocupação de grande número de habitats e o surgimento de numerosas espécies quase simultaneamente;#Evolução convergente - populações com origem diferente e sem qualquer parentesco, sujeitas a pressões selectivas semelhantes, vão desenvolver estruturas e modelos de vida semelhantes;#Evolução paralela - espécies distintas, mas com um antepassado comum, podem, independentemente, permanecer semelhantes devido ao mesmo tipo de pressões selectivas.visão criacionista de especiação
http://www.unasp-sp.edu.br/campus/neo/downloads/gibson2.pdfhttp://www.unasp-sp.edu.br/campus/neo/downloads/gibson1.pdfhttp://www.unasp-sp.edu.br/campus/neo/downloads/gibson3.pdfhttp://www.portaleducacao.com.br/forum/forum_posts.asp?TID=517&PID=1947 (ver mais especificamente em Baraminologia)Encontrei 3 linhas de argumentação que parecem fundir em torno do que tentamos defender:1.1 Há quem enfatiza homozigose como um bom fator evolutivo, apesar dos imensos riscos1.2. Outros enfatizam exatamente o contrario, os riscos e a impossibilidade de sobrevivência com populações muito pequenas , requerendo cruzamentos entre especies primas na fundação, o que gera grandes complicações1.3. Há tambem quem defende justamente poucos descendendo muitos, a exemplo da capacidade de alguns poucos casos hoje, o que recomendaria mais nosso modelo dadas as dificuldades que poderiam ser suplantadas ou minimizadas com a idéia da hipervariabilidade ancestral
Estudos 1 em populações determinam que a existência de poucos indivíduos numa população de uma determinada espécie, indica em geral baixa VG (Variabilidade genética), ou seja, podem variar-se geneticamente pouco, e uma população com poucos indivíduos têm alto grau homozigótico (características geneticamente uniformes), trazendo sérios problemas a raça pelo aumento de freqüência de compartilhamento de alelos de genes deletérios gerando a impossibilidade de sobrevivência tendo em vista a baixa VG daquela população, pois os cruzamentos em uma cessação assim aumentarão a freqüência de transmissão de alelos deletérios, onde qualquer mutação deletéria pode se espalhar e fixar rapidamente sobre aquela frágil população, o que pode em pouco tempo condená-la a extinção. Neste trabalho procuraremos ressaltar esta dificuldade de sobrevivência com o objetivo de demonstrar que somente uma configuração genética diferenciada, mais perfeita, com menor carga genética e maior patrimônio genético, na maioria das espécies, poderia permitir a explosão populacional humana e de todos os seres vivos."Existem diversas cidades do Brasil onde casamento e reprodução só são realizados entre parentes. Como exemplo: Estrela D'Oeste - SP, o número de parentes casando com parentes são extremamente altos e não há nenhum caso de doença deletéria na família" 2. Sugerimos que estas exceções de comportamento nos cruzamentos entre parentes demonstram um pouco do retrato genético ideal que existia nas populações ancestrais.Localização de Estrela do Oeste
Cap 1 ..Genealogia e Ancestralidade ComumEstudos na universidade da Yale , Dr Chang, publicado na Nature, 2004, em estatística genealógica 3, e outros mais, em busca de ancestralidade comum, como da Eva mitocôndria, de datação com base de médias de intervalos regulares de mutações, demonstram a ancestralidade comum em humanos , e o fato de termos ancestrais comuns a todos vivos e mortos, implica em avistar em uma certa época, um numero populacional reduzido, com todos os problema genéticos que hoje avistamos. Estudo feito nos Estados Unidos mostra que o mais recente ancestral comum a toda a população mundial atual, viveu, não há dezenas de milhares de anos, mas há apenas 3 mil anos . Simulações feitas em computador, baseadas em um novo modelo estatístico, mostraram que o ancestral comum a toda a humanidade não é tão velho como se imaginava. Cientistas norte-americanos descrevem na edição de 30 de setembro da revista Nature que tal indivíduo teria vivido há apenas 3 mil anos. Esse retorno com cada vez menor numero populacional nos desafia a encontrar hipóteses não uniformistas sobre a capacidade de nossos poucos ancestrais comuns nos descenderem, sem que os levasse a extinção.
Estudo feito nos Estados Unidos mostra que o mais recente ancestral comum a toda a população mundial atual, viveu, não há dezenas de milhares de anos, mas há apenas 3 mil anos . Simulações feitas em computador, baseadas em um novo modelo estatístico, mostraram que o ancestral comum a toda a humanidade não é tão velho como se imaginava. Cientistas norte-americanos descrevem na edição de 30 de setembro da revista Nature que tal indivíduo teria vivido há apenas 3 mil anos. Esse retorno com cada vez menor numero populacional nos desafia a encontrar hipóteses não uniformistas sobre a capacidade de nossos poucos ancestrais comuns nos descenderem, sem que os levasse a extinção.
1.2 Analisando Estudos sobre a Eva Mitocondrial (DNAmt)
"Os genes de uma criança são aproximadamente igualmente herdados do pai e da mãe. O rearranjo genético que ocorre a cada geração sucessiva dá ao feto duas cópias de cada gene de uma escolha de quatro possíveis dentre os genomas dos pais. A herança do DNA mitocondrial (DNAmt) é uma exceção à reprodução sexual. De singular na teoria da Eva Mitocondrial é a descoberta de que o DNAmt é herdado apenas da mãe, não havendo nenhuma contribuição do pai na época da fertilização. Quando o espermatozóide penetra o óvulo, não há entrada de mitocôndria. Apenas o DNA da cabeça do espermatozóide contribui para a formação do zigoto. A única fonte de DNAmt do zigoto é materna. A mãe passa o seu componente de DNA mitocondrial para suas filhas e filhos. Os filhos não podem passar sua informação genética adiante; a informação pode ser passada apenas pelas filhas"."O resultado surpreendente foi que todas as ramificações da árvore se juntaram em um tronco rapidamente. Rapidamente significa que usando uma taxa assumida para as mutações no DNAmt, todos os indivíduos testados poderiam ser considerados tendo uma mãe comum africana em um passado evolutivo recente. Estes investigadores concluiram: "Todos estes DNAs mitocondriais partiram de uma única mulher que se postula ter vivido 200.000 anos atrás, provavelmente na África. Todas as populações examinadas, exceto a população africana, têm origens mútiplas, implicando que cada área foi colonizada repetidamente." O ancestral africano foi chamado Eva Mitocondrial pela imprensa. A descoberta de uma convergência na árvore evolutiva a um único ancestral materno comum que viveu na África sub-Saárica há apenas 200.000 anos é sem dúvida dramática".
tenciais falhas nessa teoria têm sido extensivamente debatidas. Os 200.000 anos estimados ao ancestral comum poderiam ser um erro significante se fosse assumido que a taxa de mutação para o DNAmt estivesse errada. O DNA mitocondrial possui uma taxa de mutação mais alta do que o DNA nuclear, porque a mitocôndria não possui o mecanismo de reparo extensivo do DNA, presente no núcleo da célula". 13"Essa idade de 200 mil anos foi conseguida assumindo que o relogio molecular do DNAmt apresentasse uma taxa constante de mutação a cada 6 mil a 12 mil anos, algo em torno de 600 gerações (Gibbons, 1998). "Entretanto" escreve Wellington Silva, doutorando em Genética Humana na UNB, estudos revelaram uma taxa mutacional cerca de 20 vezes mais rápida, ou seja, uma mutação a cada 25 ou 40 gerações (Howell, 1996; Pearsons et al., 1997) . Estes dados , suscita am sérios questionamentos aos métodos utilizados para datar even6tos evolutivos (Paabo, 1996; Gibbons, 1998), enquanto para os criacionistas , soaram como boas-novas por estar em harmonia com o modelo de uma curta cronologia para nossa origem (Wieland, 1998)" 14Ou seja, voltamos a cronologia arqueologica e bíblica e encontramos novamente Adão e Eva, será mesmo mito (no sentido usual) como dizem?

Baseando-se no ADN de um antigo czar russo, os investigadores descobriram que talvez o ADN mitocondrial esteja a sofrer mutações 20 vezes mais rápido do que se pensava.
Os autores também fazem referência a um outro estudo onde os investigadores sequenciaram 610 pares de base do ADN mitocondrial de 357 indivíduos de 134 famílias diferentes e repararam que as mutações ocorrem com muito mais frequência do eles pensavam. Estudos evolutivos anteriores fizeram os investigadores pensar que iriam encontrar uma mutação a cada 600 gerações (uma a cada 12.000 anos). No entanto, ficaram "estonteados" por encontrarem alterações em 10 pares de base, o que dá uma mutação a cada 40 gerações (uma a cada 800 anos).
Mais para a frente, o artigo apresenta a controvérsia:
"researchers have calculated that "mitochondrial Eve"-the woman whose mtDNA was ancestral to that in all living people-lived 100,000 to 200,000 years ago in Africa. Using the new clock, she would be a mere 6000 years old." - (Os investigadores calcularam que a "Eva mitocondrial" - a mulher cujo mtADN foi ancestral de todos os seres humanos - viveu entre 100.000 a 200.000 anos atrás em África. Utilizando o novo relógio, ela teria uns meros 6000 anos)
Fonte 1: http://www.dnai.org/teacherguide/pdf/reference_romanovs.pdfFonte 2: http://www.nature.com/ng/journal/v15/n4/pdf/ng04
Cap 2 -Baixa Variação Genética , doenças genéticas e Perigo de extinção" Deve-se ressaltar que a variabilidade genética é a base da perpetuação de todos os seres vivos" 4 Pequenas populações apresentam em geral (não sempre) baixa VG, estas apresentam muitas doenças, fragilidades e deformidades, e está quase sempre condenada a extinção quanto maior for à taxa de cruzamentos endogâmicos"Para uma Espécie poder se reproduzir e viver em um meio é necessário um número mínimo de espécies geneticamente diferentes. Sem essa variabilidade, mudanças no meio são fatais e ainda começam aparecer problemas de deformações e doenças consideradas raras pelo seu caráter recessivo, semelhante ao ocorrido em cruzamentos com pais e filho (caso bovino) ou entre primos (com os humanos) Por isso uma espécie é considerada oficialmente extinta quando o número de indivíduos é inferior a um número mínimo, em média 25. 
Por isso animais como a ararinha azul, e a subespécie chinesa dos tigres apesar de ainda terem exemplares vivos em cativeiro e na natureza, são consideradas extintas.  Estudos em populações atuais determinam que a existência de número de indivíduos numa espécie a ameaçam com diversas doenças genéticas e extinção. ex: Variabilidade genética de peixes-bois é alarmante
Estudos em populações atuais determinam que a existência de número de indivíduos numa espécie a ameaçam com diversas doenças genéticas e extinção. ex: Variabilidade genética de peixes-bois é alarmante 
Análise de DNA mitocôndria reflete drástica redução populacional e risco de extinção 5 Se para estudos em populações, quanto menor o numero de indivíduos , maior a ameaça de extinção, pela, em geral, baixa VG ,logo, é necessário que no passado recente, quando percebemos por estatística genealógica a presença de poucos ancestrais comuns (evidenciado por estatística genealógica, relógio molecular do DNAmt, cálculos matemáticos e fontes arqueológicas diversas, para pelo menos, os humanos) , que estes indivíduos da população anterior tenham características especiais genéticas e não uniformistas, diferentes da atualidade, que suportassem e compensem as ameaças de doença e de extinção, que hoje podem ser observadas
2.1 Os Vórtices da Extinção (Extinction Vortex)
- A Classe "R" do Vórtice é iniciada quando ocorrem distúrbios que diminuem o tamanho de uma População aumentando a variação do número de indivíduos nesta. As populações ficam vulneráveis a distúrbios posteriores e uma das principais conseqüências pode ser a mudança na proporção de machos para fêmeas, o que poderia acarretar numa diminuição brusca nas taxas reprodutivas;
- A Classe "D" do Vórtice ocorre quando o tamanho da População diminui e a sua distribuição espacial fica fragmentada. Além da diminuição do fluxo gênico entre os fragmentos, o risco de extinção da espécie em fragmentos se torna maior;
- O Vórtice "F" é o aumento do número de Homozigotos devido à diminuição no tamanho da População. Há o aumento da Oscilação Genética e a conseqüente perda de alelos e variabilidade genética, causada pelo aumento de cruzamentos entre indivíduos geneticamente semelhantes (cruzamentos consangüíneos);
- O Vórtice "A" ocorre devido às mesmas causas das outras Classes de Vórtice. A diminuição do tamanho populacional resulta em menos variabilidade genética (como no vórtice "F") e consequentemente há uma redução no Potencial de Adaptação de uma população a distúrbios externos (doenças ou mudanças climáticas, por exemplo).
"Darwin nos ensina desde 1859, as espécies extintas nunca serão substituídas ou copiadas, nunca haverá um ser que consiga ser igual a uma espécie extinta. Primack e Rodrigues, 2002, consideram as espécies como "soluções para a vida no planeta" e o crescente número de extinções não nos permitirá responder questões primordiais sobre a vida e a evolução na Terra".idem 1
2.1 Os Perigos da Endogamia Continuada

"O resultado final da "Endogamia continuada (característica de populações pequenas) é a perda terminal do vigor e a provável extinção da variabilidade genética, diminuindo a fertilidade, aumentando a taxa de mortalidade e aumentando a ocorrência de anomalias genéticas" 6
"Almeida (2004), em seu artigo intitulado Teoria das Ilhas Genéticas, propõe que as raças melhoradas devam ser tratadas também como em risco de extinção, argumenta que a perda de GENES causada pela endogamia é mais perigosa que a extinção de uma raça, pois reduz a variabilidade genética e a capacidade de reação a desafios futuros, como mudanças climáticas ou mesmo novas enfermidades" 7
2. 3 Situações Ideais Genéticas para Sobrevivência das Espécies Ancestrais
Sob baixa população, para que a população sobrevivesse milênios após, teríamos que ter na espécie um "fitness" perfeito em ambiente perfeitamente adequado, alta heterose sem alelos considerados deletérios, nenhuma ou baixíssima carga genética, alta Variabilidade genética apesar da reduzida população (algo raro de se observar hoje)
Ou seja, nada que lhes ameaçasse a extinção pela transferência alélica que seria quase homogenia de caracteres deletérios. Estes deveriam praticamente inexistir.
Escreveu o biólogo Maximiliano a respeito do fato da existência do casal fundador de nossa espécie:
"Aparentemente, para contrabalançar o efeito negativo do baixo pool gênico (máximo de quatro alelos por gene, já que são dois indivíduos) um casal do tipo ancestral teria de ter:
1 -Fitiness igual ou muito próximo ao ótimo;
2 - Alta heterose (=heterozigose, = vigor híbrido), e de preferência sem alelos considerados deletérios;
3 - Baixa carga genética
4- Fatores ligados ao retardo do envelhecimento

Todavia, alguns modelos matemáticos (no livro Introdução à Genética, Lewontinl), prevêem que em poucas gerações o número de homozigotos tende a aumentar demais na população, o que com certeza é prejudicial, pois acarreta em redução do fitness devido aos cruzamentos consangüíneos próximos. Apesar de que, se a carga genética é baixa, e não encontramos alelos deletérios, a homozigose não seria tão catastrófica assim. Por outro lado, esses modelos matemáticos não parecem levar muito em consideração os efeitos da recombinação, elementos genéticos transponíveis (procurar por Ages - Altruistic Genetic Elements, na literatura criacionista) e mutações, na geração de novos alelos, então talvez os pontos a serem levados em consideração neste caso são: 1 - Se estes processos são totalmente aleatórios ou se são regulados de alguma forma (aparentemente existem evidências pra se confiar que a recombinação e a transposição, são sim bem reguladas, haja vista a complexidade dos processos); 2 - Com que rapidez esses processos ocorrem, ou seja, quanto tempo leva pra um processo desses produzir um alelo novo a ser adicionado ao pool gênico". 8Cap 3 -Aumento de Potencial de Variabilidade nos genótipos ancestrais A população de Chita têm uma variabilidade genética geralmente baixa "as crias da chita sofrem de elevados índices de mortalidade devido também a factuais genéticos" "Alguns biólogos defendem a teoria de que, em resultado da procriação consangüínea, o futuro da espécie está comprometido" . As chitas estão incluídas na lista de espécies vulneráveis da IUCN (União Internacional pela Conservação da Natureza), como subespécie africana ameaçada e subespécie asiática em situação crítica. É considerada uma espécie ameaçada de extinção no Apêndice I da CITES (Convenção sobre o comércio internacional das espécies da fauna e da flora selvagem ameaçadas de extinção)." 9
A população de Chita têm uma variabilidade genética geralmente baixa "as crias da chita sofrem de elevados índices de mortalidade devido também a factuais genéticos" "Alguns biólogos defendem a teoria de que, em resultado da procriação consangüínea, o futuro da espécie está comprometido" . As chitas estão incluídas na lista de espécies vulneráveis da IUCN (União Internacional pela Conservação da Natureza), como subespécie africana ameaçada e subespécie asiática em situação crítica. É considerada uma espécie ameaçada de extinção no Apêndice I da CITES (Convenção sobre o comércio internacional das espécies da fauna e da flora selvagem ameaçadas de extinção)." 9
"Os caçadores exibem como troféus animais diferenciados com a chita de pelo longo e as de pelo cinzento e azul indicando que no passado havia uma diversidade genética muito maior nesta espécie hoje ameaçada de extinção". idemTal tendência que chamamos de funil de expressões da variabilidade ascendente pode ser demonstrada em quase todas as espécies. Onde se percebe que espécies ancestrais possuem cada vez maior capacidade de variar-se no passado.Outro fator é que "toda vez que há um cruzamento os cromossomos se segregam aleatóriamente e se unem novamente nos gametas, também aleatóriamente. Como o novo indivíduo terá apenas 2 alelos, os outros dois ficaram de fora. Levando isso a nível de "população", por razões casuais, a cada geração teríamos uma menor variabilidade genética. Isso irá homogeneizar os indivíduosPara facilitar, analogia:Temos um dado comum, com 6 lados. Digamos que lançaríamos este dado 6 vezes em várias séries. Cada lançamento representa um indivíduo e cada série representa uma população. Toda vez que um dado número não aparecer na série de 6 ele será substituído pelo que aparecer com maior frequência.Primeira série:3 - 6 - 4 - 1 - 2 - 2 = 5 não apareceu, então mudaremos o 5 pelo 2. Agora nosso dado tem 2 faces com o número 2.Segunda série:6 - 2 - 4 - 1 - 2 - 4 = 3 não apareceu, então podemos substituir por 2 ou 4, que apareceram com maior frequência.E assim por diante. Você pode notar que toda vez que substituímos um número ele terá maior probabilidade de aparecer em detrimento dos outros. No final teremos um dado com todas as faces iguais, ou seja, a variabilidade perdida". 3.1 Análise de Progressões Populacionais ConvencionaisEis as estimativas usuais para o crescimento da população humana mundial desde a pré-história (anos negativos devem ser entendidos como a.C.):Ano população crescimento % a.a.-1.000.000 125.000 n/d-300.000 1.000.000 0,0003%-25.000 3.340.000 0,0004%-10.000 4.000.000 0,0012%-8.000 4.500.000 0,0059%-5.000 5.000.000 0,0035%-4.000 7.000.000 0,0337%-3.000 14.000.000 0,0693%-2.000 27.000.000 0,0657%-1.600 36.000.000 0,0719%-1.000 50.000.000 0,0548%-800 68.000.000 0,1539%-500 100.000.000 0,1286%-400 123.000.000 0,2072%-200 150.000.000 0,0993%1 170.000.000 0,0623%14 171.000.000 0,0451%200 190.000.000 0,0567%350 190.000.000 0,0000%400 190.000.000 0,0000%500 195.000.000 0,0260%600 200.000.000 0,0253%700 210.000.000 0,0488%800 220.000.000 0,0465%900 242.000.000 0,0954%1000 265.000.000 0,0908%1100 320.000.000 0,1888%1200 360.000.000 0,1179%1250 360.000.000 0,0000%1300 360.000.000 0,0000%1340 370.000.000 0,0685%1400 350.000.000 -0,0926%1500 425.000.000 0,1943%
1650 545.000.000 0,0000%
1700 610.000.000 0,2256%
1750 720.000.000 0,3321%
1800 900.000.000 0,4473%
1850 1.200.000.000 0,5770%
1875 1.325.000.000 0,3972%
1900 1.625.000.000 0,8197%
1920 1.813.000.000 0,5489%
1925 1.898.000.000 0,9206%
1930 1.987.000.000 0,9207%
1940 2.213.000.000 1,0831%
1950 2.516.000.000 1,2915%
1955 2.760.000.000 1,8684%
1960 3.020.000.000 1,8168%
1965 3.336.000.000 2,0103%
1970 3.698.000.000 2,0818%
1975 4.079.000.000 1,9806%
1980 4.448.000.000 1,7471%
1985 4.851.000.000 1,7497%
1990 5.292.000.000 1,7555%
1995 5.761.000.000 1,7128%
2000 6.272.000.000 1,7142%
Perguntas sobre o gráfico
-1.000.000 125.000 n/d
-300.000 1.000.000 0,0003%
-25.000 3.340.000 0,0004%
-10.000 4.000.000 0,0012%
-8.000 4.500.000 0,0059%
-5.000 5.000.000 0,0035%
-4.000 7.000.000 0,0337%
-3.000 14.000.000 0,0693%
Porque entre 4000ac e 3000ac a população dobrou de 7milhões para 14 milhões? Porque justamente entre 8000 AC e 5000 AC a população aumentou apenas 12%? De 4,5milhões para 5milhões? Ou seja, em um período de 1000 anos a população dobra, dá um salto de 7milhões pra 14 milhões, mas, em um período de três mil anos, três vezes maior que o anterior, a população aumenta míseros 12%? Ou seja, subiu de 4,5milhões para apenas 5 milhões?
Muitas respostas poderão existir, sem nenhuma base muito sólida, mais inferências conjecturais indiretas , e em franca desatenção às estatísticas genealógicas e fontes arqueológicas, bem como a uma lista de fatores que denominamos fatores de afunilamento populacional, são eles:
1.Línguas de afunilam a 3, 4 , 5 troncos básicos em 4-8 .000 AC
2.População por calculo estatístico inicia a 4-8.000 anos
3.Costumes e culturas cada vez mais semelhantes no recente passado
4.Genealogia (Yale, Nature aponta um ancestral comum a "apenas" 3000 anos)
5.Historia inicia em 4-8 mil anos
6.Número de Cidades reveladas pelas escavações
7. Referencias ao ciclo semanal de 7 dias
3. 2 Perímetro de convergência temporal
3.1.Vetor genealógico, estatísticas (Yale, dr Chang, Nature 2004) e genealogias antigas
3.2. Testes em C14 possível em rochas e diamantes(Dr Baumgardner) diminuindo tempo geológico convencional e apontando para idades recentes de camadas sedimentares e até pré-cambriana
3.3.Taxas diagênicas em fosseis sem sofrer total permineralização encontradas em ossos de T-rex com sua maior parte preservada e contendo carne com elasticidade ainda (pesquisas minhas)
3.4.Hipervariabilidade de patrimônio genético ascendente sem acumulo de doenças genéticas apontando um paraíso genético no passado sem o acúmulo delas (Pesquisas minhas semelhantes as do Dr Sanford)
3.5. Convergência de cronologias antigas
3.6. Estudo de afunilamento de nomes, de numero de cidades e referencias antigas
3.7. Criações instantâneas (Robert Gentry) com diversas publicações na Nature, Science,desbancando decaimento continuo e estabilidade "ad infinitum" de isótopos radiotivos nos métodos de datação.
Obs.:.Abundancia de Fósseis em contradição com a previsão de Darwin que seriam raros, pressupõem a sua formação catastrófica assim como a Bíblia e 272 fontes arqueológicas testificam, no caso se acoplam com este pensamento quando pensamos em catástrofes e não em histórico gradual fóssil, o modelo catastrofista explica a separação deles por camadas ao citar diversos fatores como zoneamento ecológico, (onde viviam os tipos fossilizados de baixo para cima) mobilidade, flutuabilidade de corpos, ação seletiva de águas .
Berthaut G. 1986, em "Sedimentology-experimens on lamination of sedimdiments" demonstrou em laboratório a formação SIMULTANEA de estratos tanto na vertical quanto na horizontal, revivendo e convalidando as observações de Johanes Walther, ou seja, demonstrou que a maioria das camadas geológicas formadas por ação hidrodinâmica se formaram simultaneamente, onde foi demonstrado que princípios de estratigrafia não se aplicam aos depósitos estratificados formados em canaletas de laboratório, quando há escoamento e os experimentos confirmam e explicam os estratos escalonados que Johanens Walther tinha observado na fácies estudada por ele. Eles mostraram que os Princípios da superposição e da continuidade não se aplicam em nenhum dos casos: nem a fácies nem os estratos em seqüências sucedem-se cronologicamente. Ambos foram depositados simultaneamente para formar a seqüência estratigráfica. Os experimentos confirmam a lei de Walther em fácies.
Os experimentos de Rubin e southard realizados na canaleta , juntamente com as observações submarinas, estabeleceram a relação entre as condições hidrodinâmicas e as estruturas e depósitos. Esses depósitos e estruturas são encontrados em rochas sedimentares, e a partir delas podem ser determinadas às condições hidrodinâmicas, em particular a velocidade do escoamento, como Austin fez com relação ao Grupo Tonto. O valioso trabalho dele foi complementado com a descoberta do mecanismo de formação dos estratos, em nossos experimentos. 16.
3.3 Análise de Parâmetros para Simulações de Sobrevivência de Espécies AncestraisTentatiivas de simular como números extremamente baixos de população de espécies como as citadas pela Bíblia, tem sido feitas, mas sem respeitar todos os itens , ou pelo menos o que se pode avistar deles com progressões de como seria o DNA de 4000 anos atrás. Em simulações assim o resultado é sempre negativo, ou seja, em poucas gerações, as espécies se extinguem. Em debate sobre este tema com pesquisadores da área fui informado:"Mesmo considerando a taxa de mortalidade como zero, o crescimento populacional chega ao máximo a 5,73% . Como é que 8 indivíduos conseguiriam chegar aos 6 bilhões atuais? (sem contar que depois de tantas guerras, desastres e epidemias tivemos grandes "baixas" nesse número)??? Esquece as mutações, pense somente numa população pequena e isolada sofrendo deriva genética...Eu fiz a simulação para apenas 10 loci genênicos, todos inicialmente heterozigotos (variabilidade genética inicial 100%, como sugere o modelo bíblico), e os resultados foram: - Não sobrou nenhum heterozigoto (freqüência = 0.5)Ou seja, na primeira mudança no ambiente, na primeira pressão seletiva que houvesse, e a população se extinguiria".17 Conforme o quadro abaixo:
Outro desafio que fazemos ao modelo evolucionista é que demonstrem como as mais de 500 espécies, supostamente "bem adaptadas", que atravessaram "supostamente" milhões de anos, muitas desde o cambriano (considerado como tendo existido a 500 milhões de anos atrás) estão em flagrante extinção se foram elas, semelhanes genóptica e fonópticamente, que fundaram e sobreviveram a tragédias e isolamentos por tanto tempo?
O comportamento cientifico atual, em relação à sobrevivência de espécies ancestrais sobre os moldes darwinistas, se apresentam bastantes distantes de uma síntese, justamente por assumirem um certo atualismo genético, que torna nossa ancstralidade genealogicamente falando incapaz de sobreviver e dar bilhões de descendentes atuais.
Por um lado defendem a sobrevivência até em homozigose, destacando que isso ajudaria evolução e surgimento de novas espécies, pouco dando atenção ao aspecto sobrevivência. Por outro lado, defendem exatamente o oposto quando se apresenta um modelo de poucos ancestrais como o bíblico, com espécies hipervariáveis, saudáveis e longevas, neste caso partem para demonstrar com gráficos a extinção mesmo até em heterozigose máxima (claro que não levando em conta diversos aspectos)
A solução mais sensata a meu ver seria uma ancestralidade diferente, mais longeva, mais hipervariável, com acumulo quase zero de mutações deletérias, assim como podemos progressionar em diversas espécies como a "cheetah", onde caçadores ainda exibem a variação "azul" de algumas décadas atrás. Contudo, o preconceito anti-religioso não permitiria tal linha de raciocínio em meio acadêmico, e se criaria muito alvoroço e protestos, claros e escondidos
O certo é que os parâmetros que poderíamos deduzir para um DNA ancestral dentro desta perspectiva bíblica não mais existem, devido a muitas deleções de genes , genes que se perderam na quase eliminação das espécies durante o dilúvio, patrimônio genético perdido, genes não foram selecionados e fixados, mutações que não existiam não se pode monitorar, etc.Então que parâmetros teríamos?"
Testar um modelo sem estas e muitas outras informações, tais como informações de longevidade, seria impossível. Talvez se perseguir o máximo de dados disponíveis em raras especies de baixas populações e com alta VG, ou tanto no relato de Gênesis como na ciência moderna que os espelham, tais como milhares de fatores genéticos ligados ao envelhecimento, seria uma saída mais razoável, mas não menos complexa.
O aumento de hipervariabilidade genética claramente vista nas espécies ancestrais, deveria ser fator importante nesta averiguação. Pois temos casos hoje de poucos indivíduos conseguindo sobreviver e deixar muitos descendentes, bem como casos raros de populações pequenas com alta VG. A pergunta que nos surge é: Quê lições estes casos nos ensinariam? Até que ponto espelha o que pôde ter acontecido no recente passado?
Para que se façam tais simulação é fundamental que enxerguemos que a necessidade de heterozigose hoje não é a mesma, pois que os cruzamentos "salvadores" de espécies ameaçadas que se fazem hoje, acontecem com espécies primas, ou seja, espécies que descenderam de espécies-mães comuns, apontando para um maior patrimônio e potencial anterior, sempre que retornamos
Ora, não precisa calcular muito para entender que se retornarmos míseros 4000 anos encontraremos DNAs hipervariáveis, sem mutações deletérias e excepcionaisUm exemplo do que falamos em historia antiga é a quase ausência de relatos de natimortos apesar de todas as culturas incentivarem casamentos primos, o que até pelas baixas populações era algo forçoso.http://www.orkut.com/Profile.aspx?uid=60408417562233448313.4 Justificativas que Demonstram Longevidade nas Populações AncestraisApesar de muitos defenderem aumento de expectativa de vida da população atual, lembramos que "o que a medicina aumentou foi a expectativa de vida e não a idade máxima que as pessoas alcançam. Expectativa de vida é a média das idades. Se você combate a mortalidade infantil você aumenta a expectativa de vida, a idade média das pessoas mas eventualmente não acrescenta nada às suas idades máximas""É interessante observar que as idades máximas registradas foram de escravos e pessoas do terceiro mundo. O que o primeiro mundo registrou foi um percentual maior de pessoas idosas mas os recordes de idade não estão com eles. Expectativa de vida se relaciona com as doenças e o combate delas. Quanto maior a densidade demográfica e ou menor o grau de higiene, menor a expectativa de vida útil" 18
Outros fatores ligados a longevidade são:
3.4.1. Jacks Cuozzo prova hominideos vivendo mais 300 anos
http://www.jackcuozzo.com/swanscombe.html
3.4.2. Padrão fossil maior dos seres vivos , possuem estreita ligação com maior longevidade
3.4.3. Comparação clara de pessoas que vivem em lugares mais naturais , com habitos frugais, e mais longevidade..em toda parte
3.4.4. Tenho evidencia de alimentação mais vegetariana de neandertais pelos dentes (Cuozzo e outros), e evidencia de longevidade em cidades de população de vegetarianos como Loma Linda
http://br.youtube.com/watch?v=2PYfrvaQ36w
3.4.5. Engana-se quem pensa que na antigiudade o homem era "carnívoro", ou comia predominantemente carne. Estudos arqueológicos de Burkit concluiram que o homem consumia cerca de 300g de fibras por dia (que só existem em vegetais). Atualmente o brasileiro médio consome em média 13g de fibras diariamente. AS recomendações de fibras estão entre 25-50g (dependendo de idade e sexo)". Pouca fibra=câncer, e vegetarianismo = maior longevidade, ESTÁ PROVADO SUYN, I'm sorry pelos evos, mas a ciência agradece
3.4.6. A Biblia, que é documento arqueológico, é uma evidencia que se demonstra honesta pois não esconde até mesmo erros do seu proprio povo, ao contrario de muitas fontes
3.4. 7. Centenas de relatos de outros povos, onde a tradição de falar de longevidade ancestral é uma marca
3.4.8. Progressões de aumento/diminuição de acúmulo de mutações deletérias em pesquisa de 5000 doenças genéticas (Dr Sanford) o que implica diretamente em longevidade dos ancestrais
| |
|
Cap. 4. Conclusão
Concluímos que a única saída para se explicar a não extinção das espécies ancestrais é que as mesmas possuíam genética diferenciada, algo próximo ao que a Bíblia relata. Que as espécies atuais, na sua maioria, já desgastadas, tendo perdido volume patrimonial genético e feitas às adaptações das mais diversas para sobreviverem, estão por sua vez, cada vez mais comprometidas e frágeis, como se demonstra hoje em inúmeras extinções, mesmo com posterior protecionismo humano. O que percebemos hoje, é que caso espécies sofram algumas pressões ambientais, elas se extinguem, devido a vários fatores e principalmente ao acumulo de carga genética que a seleção natural foi e é incapaz de conter (Dr Sanford), e principalmente por terem fugido de sua configuração original, a qual lhes permitia suportar mais alguma sobrevida, pois que estariam cada vez mais próximas do plano original do Criador.
Sofremos em media por dia 500.000 reparos em nosso DNA! Isso se dá devido a força fraca que unem as letras do DNA das células, esta realidade nos fala de um código genético fragil e ameaçado em toda historia humana e dos seres vivos. Nos fala de antepassados com menor suceptibilidade de defeitos que os seres atuais, o que nos recomenda a não só dar um basta na direção de melhora dos caminhos evolutivos sobreviventes, como já fez o neutralismo, mas de reinvindicar a tendencia entrópica para o futuro, o que nos recomenda automaticamente para um paraíso perdido no recente passado.
4 RECH, Elíbio "A biotecnologia, a sustentabilidade e a biodiversidade Brasileira" "Qual o impacto da biotecnologia sobre a diversidade genética?" Disponivel em <http://www.cib.org.br/pdf/biotecnologia_sustentabilidade_biodiversidade%20.pdf > <http://www.cib.org.br/faq.php?id=4> . Acesso em 19 de maio de 2008 5 KUCK, .Denis Weisz em "Variabilidade genética de peixes-bois é alarmanteAnálise de DNA mitocondrial reflete drástica redução populacional e risco de extinção" Disponivel em <http://cienciahoje.uol.com.br/controlPanel/materia/view/2646> acesso em 19 de amio de 20086. FALCÃO, a j s; "Efeitos da Endogamia sobre Características de Reprodução, Crescimento e ValoresGenéticos Aditivos de Bovinos da Raça Pardo-Suíça" ROBERTSON, A. 1961. Inbreeding in artificial selection programmes. Genet. Res., 2:189-194 . Disponivel em <http://64.233.183.104/search?q=cache:rUIw4YQVQqEJ:www.scielo.br/pdf/rbz/v30n1/5439.pdf+%22Os+pr%C3%B3s+e+contras+da+endogamia%22&hl=pt-BR&ct=clnk&cd=2&gl=br> Acesso em 19 demaio de20087. .ALMEIDA, m J Oliveira; "CONSERVAÇÃO DE RECURSOS GENÉTICOS:, UTILITARISMO, PRESERVACIONISMO E CONSERVACIONISMO" Disponivel em < http://64.233.183.104/search?q=cache:TnCrKfR62i0J:www.ruralnet.com.br/upload/artigos/CONSERVA%C3%87%C3%83O%2520DE%2520RECURSOS%2520GEN%C3%89TICOS.doc+extin%C3%A7%C3%A3o+popula%C3%A7%C3%A3o+variabilidade+gen%C3%A9tica&hl=pt-BR&ct=clnk&cd=1&gl=br> Acesso em 19 de amio de 20088. MENDES, maximiliano em "Problema Dificil de Resolver" Disponivel em <http://www.orkut.com/CommMsgs.aspx?cmm=5266336&tid=2439440642721406082&kw=pool+genico&na=4&nst=-8&nid=5266336-2439440642721406082-2440161943824063618> Acesso em 19 de maio 20089.WIKIPEDIA "Chita (animal)"Disponivel em <http://pt.wikipedia.org/wiki/Chita_(animal)> Acesso em 19 de maio de 2008
10. HARTWELL, Sarah "Inbreeding de A à Z .Prós e contras da endogamia" 2003 11. SANFORD, john " Genetic Entropy & the Mystery of the Genome"(October 2005) Disponivel em <http://en.wikipedia.org/wiki/John_C._Sanford><http://www.answersingenesis.org/PublicStore/product/Genetic-Entropy-the-Mystery-of-the-Genome,4632,226.aspx>"O Dr. John Sanford, phd,"a maioria das mutações aleatórias são deletérias, e não neutras como proposto pela TE, "seleção natural não suficiente para filtrar estas mutações, a médio e longo prazo isto levará extinção<http://www.uncommondescent.com/archives/1173>12.HARTWELL, Sarah "Inbreeding de A à Z .Prós e contras da endogamia" 2003 13. TRIUNFOL, Marcia; PARIS, M B Rivero "O DNA mitocondrial de Eva" Disponivel em <http://www.odnavaiaescola.com/artigo-eva.htm> Retirado do site O DNA vai à Escola. Acesso em 19 de maio de 200814 SILVA, W Santos, "Uma discussão sobre árvores evolutivas humanas obtidas de estudos com marcadores moleculares" Revista Criacionista numero 69 , segundo semestre de 2003, pag 3615. DELONG, B; COHEN, J E "Estimating World GDP, One Million B.C. - Present" Disponivel em <http://www.j-bradford-delong.net/TCEH/1998_Draft/World_GDP/Estimating_World_GDP.html> Acesso em 19 de maio de 200816. Berthaut G. 1986, Sedimentology-experimens on lamination of sedimdiments" Austin, S A ,1994 "Grand Canyon, Monument to catastrophe" ICR, USA < http://qao.sites.uol.com.br/apostilageseao.htm> Acesso em 19 de maio de 2008
17 SUYNDARA, "Réplica ao Sodré" Disponivel em <http://www.orkut.com/CommMsgs.aspx?cmm=26688928&tid=2597695239025054347&na=4&nst=160&nid=26688928-2597695239025054347-5202378522903958155> Acesso em 19 de maio de 200818 JANSEN, w " Adão, Seus Filhos, População Mundial e Longevidade" Disponivel em <http://www.orkut.com/CommMsgs.aspx?cmm=26796515&tid=2524926147072319710&kw=120+anos> Acesso em 19 de maio de 200819. CUOZZO,Jacks "Buried Alive: The Startling Truth about Neanderthal Man" Disponível em <http://www.jackcuozzo.com/swanscombe.html> acesso em maio de 200820. JANSEN, w " Adão, Seus Filhos, População Mundial e Longevidade" Disponivel em <http://www.orkut.com/CommMsgs.aspx?cmm=26796515&tid=2524926147072319710&kw=120+anos> Acesso em 19 de maio de 200821. ´PADRÃO, A P "SBT Realidade - Estilo de Vida Adventista" Disponivel em <http://br.youtube.com/watch?v=2PYfrvaQ36w> acesso em 19 de maio de 200822. ANDRADE, S. C "Carne e Câncer" Disponível em < http://www.orkut.com/CommMsgs.aspx?cmm=26688928&tid=2595900350638544949&kw=+Burkit+concluiram+&na=3&nid=26688928-2595900350638544949-2596186286021636227&nst=71> Acesso em 19 de maio de 2008
23 PAULO J. O Mito do 1% Disponivel em acesso em 19 de maiode 2008Um exame mais cuidado dos dados publicados provenientes do projeto do genoma do chimpanzé revela outras diferenças profundas que fazem com que a alegação de 1% de diferenças pareça um tanto tolice. Estas incluem: 1. Diferenças estruturais básicas entre os cromossomas das duas espécies incluindo o número de cromossomas, a posição dos centrómeros [3] e o número de inversões de segmentos e de relocações; 2. Diferenças profundas entre as espécies na seqüência do DNA dentro dos telomeros [4] nas extremidades de cada cromossoma; 3. Seqüências de DNA únicas em cada espécie envolvidas no controle particular e complexo de genes similares; e 4. Regiões radicalmente diferentes e especificas de cada espécie envolvidas na Recombinação Genetica [5], um processo iniciando na meiose que conduz à formação do esperma e do ovo.Um trabalho mais recente destacou diferenças entre os seres humanos e os chimpanzés que são muito mais profundas do que se tinha reconhecido anteriormente. Por exemplo, Oldham at al. publicaram um trabalho que descreve redes geneticas nos cérebros do ser humano e do chimpanzé no "Proceedings of the National Academy of Sciences" [6]. De acordo com estes autores, 17.4% das conexões de rede no cérebro foram encontradas no ser humano mas não no chimpanzé. Para além disso, Matthew Hahn e colegas de trabalho relataram uma diferença de 6.4% no número da cópias de gene entre o ser humano e o chimpanzé, num artigo de dezembro 2006 da PLoS One [7]. Com a publicação continuada de dados do projeto ENCODE, tornar-se-á cada vez mais difícil manter a mitologia do 1% de diferença entre as duas espécies. As implicações para a evolução Darwinista são muitas.Links:[1] - Science - http://www.ncbi.nlm.nih.gov/sites/entrez?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=17600195[2]- Genoma do Chimpanzé - http://www.ncbi.nlm.nih.gov/sites/entrez?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=16136131[3] - Centromeros - http://en.wikipedia.org/wiki/Centromere[4] - Telomeros - http://en.wikipedia.org/wiki/Telomere[5] - Recombinação genética - http://en.wikipedia.org/wiki/Genetic_recombination[6] - Proceedings of the National Academy of Sciences - http://www.ncbi.nlm.nih.gov/sites/entrez?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=17101986[7] - PLoS One - http://www.ncbi.nlm.nih.gov/sites/entrez?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=17183716http://www.orkut.com/CommMsgs.aspx?cmm=14251159&tid=2544078963181814212&start=1
Mais Consultas
APREMAVI (Associação de Preservação do Meio Ambiente do Alto Vale do Itajaí). No Jardim das Florestas Org. Miriam Prochnow. Rio do Sul: Apremavi, 2007.
BIRDLIFE INTERNATIONAL. Threatened birds of the world. Lynx Edicions, Barcelona, e BirdLife International, Cambridge, Reino Unido. 2000.
BLOMQVIST, D.; PAULINY, A. Caught in extinction Vortex? A Population decline and genetics in a. metapopulation of Southern Dunlins (Calidris alpine). Departament of Zoology, University of Gothenburg, Sweden. 2007.
CORDEIRO, P.H.C. A Fragmentação da Mata Atlântica no Sul da Bahia e suas implicações na conservação de Psitacídeos. Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil. 2003.
DARWIN, C. A Origem das Espécies. Texto Integral. Tradução: John Green. São Paulo: Editora Martin Claret, 629 p. 2006.
FRANÇA, V.B. Dados preliminares sobre o estudo das Epífitas (Bromeliaceae, Orchidaceae e Cactaceae) que ocorrem na reserva do Parque do Zizo - São Miguel Arcanjo, São Paulo. Centro Universitário Nossa Senhora do Patrocínio - CEUNSP. 2001.
GILPIN, M.; SOULÉ, M,E. Minimum Viable Populations: Processes of Species Extinction. pp. 19-34 in M. E. Soulé, ed. Conservation Biology: The Science of Scarcity and Diversity. Sinauer, Sunderland, Mass. 1986.
LEAL, I,R. DA SILVA, J.M.C.; TABARELLI, M. LACHER JUNIOR, T.E. Mudando o curso da conservação da biodiversidade na Caatinga do Nordeste do Brasil. Revista Megadiversidade. Vol. 1, n. 1. 2005.
MYERS, N.; MITTERMEYER, R.A.; FONSECA, G.A.B.; KENT, J. Biodiversity hotspots for conservation priorities. Nature 403: 853-858. 2000.
PRIMACK, B.R; RODRIGUES, E. Biologia da Conservação. Londrina, Paraná: Editora Planta, 2002.
RICKLEFS, R.E. A Economia da natureza. 5ª edição. Rio de Janeiro: Guanabara Koogan. 2003.
RODRIGUES, F.H.G. Biologia e Conservação do Lobo-guará Na Estação Ecológica de Águas Emendadas, DF. Tese de Doutorado. Campinas, 2002.
SOUZA, V.C. LORENZI, H. Botânica Sistemática, Guia Ilustrado para identificação das famílias de Angiospermas da flora Brasileira, baseado em AGP II. Nova Odessa: Instituto Plantarium, 635 p. 2005.
FAGAN, W.F.; HOLMES, E.E. Quantifying the extinction vortex. Ecology Letters, Vol. 9 p. 51-50. 2006
HARRIS, R.B.; MAGUIRE, L.A. SHAFFER, M.L. Sample Sizes for Minimum Viable Population Estimation Conservation Biology, Vol. 1, No. 1 , pp. 72-76. 1987
|
O declínio genético do Homem
No ano de 1993, Lori Oliwenstein escreveu na revista Discover:
Começando pela primeira célula se aglutinou dando origem à sopa primordial, até à complexidade do Homo sapiens, a evolução da vida – como toda a gente sabe – tem sido impulsionada rumo a uma maior complexidade. O único problema com o que toda a gente sabe . . é que não há a mínima evidência de que é verdade.1
Não há qualquer tipo de evidência que confirme as noções evolutivas da humanidade ir ficando cada vez mais apta. Da mesma forma que não se escapa da morte, não se escapa do atrito biológico. Os fósseis, a genética, e a História apontam para um declínio implacável da humanidade.
Os fósseis de ossos humanos revelam que os homens eram muitos maiores no passado do que são agora. Os Neandertais eram um povo antigo que vivia nas cavernas numa área que se estendia da Europa a Israel.
Um característica peculiar dos Neandertais é exagerada dimensão dos seus troncos e dos ossos dos membros. Todos os ossos preservados sugerem uma força raramente atingida pelo homem moderno.2
O antropólogo Peter McAllister pesquisou pegadas de fósseis humanos na Austrália e apurou que quem quer que fez essas pegadas corria mais rapidamente que o sprintercampeão Usain Bolt.3 Paralelamente, locais europeus pertencentes ao Paleolítico mostram que os humanos eram maiores então, e que nós experimentamos um “vincado declínio na altura“, declínio esse que nunca mais nos permitiu recuperar a altura inicial.4
O declínio no tamanho e na força parecem consistentes com o declínio genético geral observado tanto nos estudos das sequências genéticas 5 como também no programa que monitoriza as mutações chamado de “Mendel’s Accountant”. As mutações alteram a complicada informação em código celular, e a maior parte das alterações não têm qualquer tipo de efeito nas células.
Cada uma das pequenas alterações tem um impacto tão reduzido que o mesmo não é detectado ou removido pelo mecanismo de reparação genético. Simulações que usam parâmetros biologicamente realistas, tal como 60 mutações e seis filhos por cada geração, claramente mostram que a aptidão diminui a cada geração. 6
A implacável acumulação destas corrupções genéticas progressivamente corrói o código genético original:
Se as mutações sem utilidade (ou deleteriosas) são transmitidas para a geração seguinte, elas formarão de modo incremental um largo lastro de material sem utilidade.7
Os escritos antigos relatam eventos em torno de heróis com grande força física. O Épico de Gilgamesh descreve o genuíno Rei Gilgamesh como alguém extraordinariamente forte, tendo lutado com uma besta-fera com a aparência dum dragão. Muitas outras lendas contam histórias de homens fisicamente bem construídos. Beowulf “encontra-se bem registado na História. Ele foi o filho de Ecgtheow, havendo nascido no ano AD 495”, e era famoso pelo seu tamanho, que lhe permitiu lutar contra répteis gigantes que se encontram agora extintos na Dinamarca.8
As Escrituras descrevem também pessoas que eram mais altas e mais fortes que as actuais. Golias tinha mais de 2,7 metros de altura, e a sua “cota de malha” pesava cerca de 56 quilos, para além de outro equipamento de guerra que ele carregava.9 Para além disso, os Israelitas mataram Ogue de Basã, que tinha cerca de 4 metros de altura cuja cama tinha cerca de 4 metros de altura e 1,75 de largura.10
A expectativa de vida também diminuiu drasticamente, tal como registado em Génesis 11, o que, mais uma vez, revela mais perda biológica e não ganho. Este ciclo de deterioração inevitável teve início quando o pecado e a morte entraram no mundo.
Felizmente, a Bíblia proclama que o Senhor Jesus conquistou a morte e disponibiliza a redenção do pecado. A salvação vem mediante fé de que “Cristo morreu pelos nossos pecados, segundo as Escrituras, E que foi sepultado, e que ressuscitou ao terceiro dia, segundo as Escrituras,”11 Para aqueles que acreditam, o Senhor Jesus Cristo por fim irá “Que transformará o nosso corpo abatido, para ser conforme o Seu Corpo Glorioso, segundo o seu eficaz poder de sujeitar, também, a si todas as coisas”12
Referências
-
Oliwenstein, L. 1993. Onward and Upward? Discover. June: 22.
-
Trinkaus, E. 1978. Hard Times Among the Neanderthals. Natural History. 87 (10): 58. Quoted in Lubenow, M. 2004. Bones of Contention. Grand Rapids, MI: Baker, 78.
-
-
Holt, B. M. and V. Formicola. 2008. Hunters of the Ice Age: The biology of Upper Paleolithic people.American Journal of Physical Anthropology. 137 (47): 70-99.
-
See Thomas, B. The Human Mutation Clock Is Ticking. ICR News. Posted on icr.org July 7, 2011, accessed December 15, 2011.
-
In this simulation program, “fitness” directly correlates to net genetic information loss or gain accrued over a set number of generations. See Vardiman, L. 2008. The “Fatal Flaws” of Darwinian Theory. Acts & Facts. 37 (7): 6.
-
Hobrink, B. 2011. Modern Science in the Bible. New York: Howard Books, 140.
-
Cooper, B. 1994. After the Flood. Chichester, UK: New Wine Press, 148.
-
-
-
-







 Fonte
Fonte
